

De polyvagaaltheorie: huidige stand van zaken, klinische toepassingen en toekomstperspectieven (1)
Geschreven door Stephen Porges


Inleiding
Recent verscheen een artikel van de hand van Stephen Porges, grondlegger van de polyvagaaltheorie (PVT), over de huidige stand van zaken rond zijn theorie. Natuurlijk was ik direct geïnteresseerd en heb ik het artikel gedownload en doorgenomen. Ik werd enthousiast en vond het de moeite waard om integraal te vertalen. Het artikel is bijna 15 inhoudelijke pagina’s lang, in compact geschreven medisch-technisch Engels1.
Het vertalen was (dus) een hele klus. Gezien de lengte publiceer ik de vertaling in meerdere delen. Dit is het eerste deel van drie. (Terwijl ik dit schrijf, ben ik nog niet klaar met het resterende gedeelte, maar dat komt wel goed hoor.)
In zijn artikel gaat Porges in op …
de geschiedenis van de PVT,
de wetenschappelijke basis vanuit de neurofysiologie en de evolutieleer,
de rol van de ademfrequentie en de respiratoire sinusaritmie in relatie tot de neurofysiologische basis van de PVT.
Deze punten komen in dit eerste deel van de vertaling aan de orde. Dat is wat mij betreft de moeite al meer dan waard, onder meer omdat Porges hier ingaat op de argumenten die critici gebruiken om te betogen dat de polyvagaaltheorie niet deugt, en deze argumenten weerlegt.
In de volgende delen komen deze onderwerpen nader aan de orde:
de popularisering van de PVT en problemen die daaruit voortkwamen,
falsifieerbaarheid en de voorspellende kracht van de PVT,
klinische toepassing in verschillende disciplines,
methodologische innovaties en het meten van autonome toestand,
toekomstige mogelijkheden en perspectieven in de wetenschap van het autonome zenuwstelsel,
polyvagaal geïnformeerde organisaties en instituties.
Kortom, óók de moeite waard!
Geen eenvoudige kost
Zoals je hopelijk met me eens bent, staan hierboven een aantal belangrijke onderwerpen, waarover de laatste jaren, naast veel bijval en belangstelling, ook regelmatig discussie en misverstanden zijn ontstaan.
Omdat ik het artikel integraal vertaal en op dit moment verder niet voorzie van extra uitleg, zal het niet voor iedereen eenvoudig te lezen zijn. Dus als je na een paar alinea’s denkt dat dit boven je pet gaat, dan is dat niet vreemd. Ik ben zelf goed bekend met de medisch-technische taal waarin het artikel is geschreven en ik heb ook vaak moeten puzzelen en overdenken.
Dus wees gerust én verheug je: op RelaxMore.net vind je genoeg comtent die ook voor leken toegankelijk is.
Als je geïnteresseerd bent in de polyvagaaltheorie, volg dan ook het Polyvagaalinstituut Nederland.
Info en rechten
Auteur van het oorspronkelijke artikel: Stephen W. Porges
Bron: Clinical Neuropsychiatry (2025) 22(3): 169–184.
DOI: doi.org/10.36131/cnfioritieditore20250301
Licentie: CC BY-NC-SA 4.0 — Dit is een vertaling van het Engelstalige open‑accessartikel, gemaakt door Ronald de Caluwé.
Deze vertaling is niet‑commercieel en valt onder dezelfde licentie2.
De polyvagaaltheorie: huidige stand van zaken, klinische toepassingen en toekomstperspectieven
Stephen W. Porges
Samenvatting
De polyvagaaltheorie biedt een evolutionair geïnformeerd neurofysiologisch raamwerk om te begrijpen hoe het autonome zenuwstelsel sociale betrokkenheid, emotionele veerkracht en adaptieve fysiologische responsen ondersteunt. In de kern benadrukt de theorie een hiërarchische organisatie van autonome toestanden die worden bestuurd door de nervus vagus, waarbij de unieke rol van het ventraal vagale complex in het faciliteren van sociaal gedrag en fysiologische flexibiliteit via mechanismen zoals neuroceptie, co‑regulatie en dissolutie wordt uitgelicht. Dit artikel bespreekt de empirische fundamenten van de polyvagaaltheorie, gaat in op methodologische kritiek – in het bijzonder met betrekking tot anatomische specificiteit en respiratoire sinusaritmie (RSA) – en bundelt recente ontwikkelingen in autonome meetmethoden. Verder verkent het brede klinische implicaties met betrekking tot uiteenlopende aandoeningen, waaronder trauma, chronische pijn, autisme, ontwikkelings- en stemmingsstoornissen. Tot slot pleit het artikel voor toepassing van een ‘wetenschap van veiligheid’ in de klinische praktijk, het onderwijs en de publieke gezondheidszorg, met voorstellen voor toekomstig onderzoek, praktische inrichting en de systemische inrichting van instellingen die fysiologische veiligheid zouden moeten ondersteunen.
Trefwoorden: polyvagaaltheorie, ventraal vagaal complex, neuroceptie, vagale efficiëntie, trauma, autonoom zenuwstelsel
Stephen Porges
Traumatic Stress Research Consortium, Kinsey Institute, Indiana University3.
1. Inleiding: van theorie naar perspectief
Toen ik de polyvagaaltheorie in 1994 voor het eerst introduceerde, was mijn doel een neurofysiologisch model te bieden dat kon verklaren hoe het autonome zenuwstelsel – en in het bijzonder de nervus vagus – onze capaciteit voor emotie, gedrag en sociale verbinding vormgeeft. Wat begon als een wetenschappelijke poging om de lichaamsrespons op uitdaging te begrijpen, is uitgegroeid tot een breder perspectief op hoe veiligheid, regulatie en relaties in onze biologie zijn ingebed.
In de afgelopen drie decennia is deze theorie niet alleen door onderzoekers en clinici omarmd, maar ook door docenten, gezondheidsprofessionals, trauma‑overlevenden en pleitbezorgers voor sociale rechtvaardigheid. De polyvagaaltheorie groeit zowel in empirische onderbouwing als in transdisciplinaire relevantie en biedt een taal om te beschrijven hoe ons zenuwstelsel navigeert tussen toestanden van verdediging en verbinding.
De kern van de theorie is een eenvoudig, maar transformerend idee: de mogelijkheid om ons veilig te voelen en met anderen te verbinden is biologisch verankerd in de structuur en functie van ons autonome zenuwstelsel. Wanneer we gedrag bekijken door de lens van de neurofysiologische toestanden, verandert ons perspectief van oordeel naar nieuwsgierigheid, van pathologie naar adaptatie.
In dit artikel reflecteer ik op de huidige stand van zaken van de polyvagaaltheorie, haar neuro‑anatomische fundamenten en haar steeds breder wordende klinische en maatschappelijke toepassingen. Ik reageer op kritieken en misinterpretaties die zijn ontstaan en schets een toekomstgerichte visie over waar de theorie de wetenschap, de gezondheidszorg en de politiek kan ondersteunen. Voortbouwend op Polyvagal Perspectives (2024) zet dit artikel de inspanning voort om een ‘wetenschap van veiligheid’ te formuleren – een die zowel individueel welbevinden als collectieve transformatie kan begeleiden. De volgende paragrafen behandelen de wetenschappelijke basis van de theorie, haar evolutionaire- en ontwikkelingsoorsprong en de onderscheidende kenmerken van de autonome regulatie bij zoogdieren.
2. Wetenschappelijke fundamenten: neurofysiologie en evolutie
2.1 Ontwikkelings‑ en fylogenetisch bewijs
Het ontwikkelings‑ en evolutionaire traject van het ventraal vagale complex vormt de kern voor haar rol in het sociale betrokkenheidssysteem. Het ventraal vagale complex is niet slechts een ‘nieuwere’ laag anatomie – het is een evolutionaire herbestemming van oude hersenstamcircuits (Porges, 2001). Tijdens de evolutie van gewervelden migreerden cardio‑inhibitoire neuronen ventraal, van de dorsale motorische nucleus van de vagus (DMNX) naar de nucleus ambiguus (NAmb) (Porges, 2023). Deze migratie, uniek voor zoogdieren, maakte de ontwikkeling mogelijk van een gemyeliniseerde vagus die de dynamische regulatie ondersteunt die nodig is voor zogen, vocalisatie en zorggedrag (Porges, 2023). Deze anatomische veranderingen lopen parallel met de embryologische ontwikkeling, waarin de craniale zenuwen die betrokken zijn bij het sociale betrokkenheidssysteem – V, VII, IX, X en XI – ontstaan uit de faryngeale bogen (Moore et al., 2018). Hoewel ze worden vertegenwoordigd door afzonderlijke motorkernen, ondersteunt hun gedeelde ontwikkelingsoorsprong hun geïntegreerde functie in gelaatsuitdrukking, stemmodulatie, ingestie en oriëntatiegedrag (Porges, 2023).
Bij zoogdieren worden deze neurale structuren gecoördineerd door het ventraal vagale complex (VVC) om toestandsafhankelijk sociaal gedrag te reguleren, zoals zogen, hechting, vocale communicatie en prosodie (Porges, 2007). Dit systeem functioneert bij de geboorte en maakt de coördinatie van zuigen‑slikken‑ademen mogelijk en vormt het fysiologische substraat voor socialiteit en emotionele co-regulatie (Porges & Furman, 2011).
Deze evolutionaire aanpassing benadrukt de rol van de nervus vagus voor het creëren van de basis voor sociale en emotionele regulatie, waarbij fysiologische responsen verbonden zijn met gedragingen van sociale betrokkenheid (Porges, 2003). De neurale koppeling van deze systemen faciliteert dynamische interacties die essentieel zijn voor de overleving en ontwikkeling van een zuigeling en ondersteunt de integratie van autonome en sociale gedragingen zoals dat bij zich ontwikkelende zoogdieren plaatsvindt (Porges, 2007). Voortbouwend op deze ontwikkelingsbasis, vergelijk ik in de volgende paragraaf de neurale architecturen van zoogdieren en reptielen die relevant zijn voor autonome regulatie en sociale betrokkenheid.
2.2 Vergelijkende neuro‑anatomie, autonome integratie en het sociale betrokkenheidssysteem
Ter verdere verduidelijking van de uniciteit van de autonome regulatie bij zoogdieren presenteert deze paragraaf een vergelijkende analyse van de neurale structuren die ten grondslag liggen aan autonome en sociale betrokkenheidssystemen bij zoogdieren en reptielen. Critici van de polyvagaaltheorie (bijv. Grossman & Taylor, 2007; Grossman, 2023; Taylor et al., 2022) verwarren vaak anatomisch homologe structuren met functionele gelijkwaardigheid, ten onrechte aannemend dat de aanwezigheid van een NAmb bij reptielen autonome capaciteiten impliceert die vergelijkbaar zijn met die bij zoogdieren. Dit duidt op een verkeerd begrip van de vergelijkende neuro‑anatomie, die onderscheid maakt tussen homologe structuren – met een gedeelde evolutionaire oorsprong – en analoge functies, die onafhankelijk kunnen ontstaan in onderscheiden lijnen. De redenering van Grossman en Taylor veronderstelt onterecht dat gedeelde kenmerken, zoals cardio‑inhibitoire vagale banen, gedeelde functie impliceert, en ziet de evolutionaire reorganisatie en diversificatie van deze systemen over het hoofd.
De polyvagaaltheorie daagt deze aanname uit door te stellen dat de evolutie bestaande anatomische substraten herbestemt voor nieuw ontstane functies. Hoewel reptielen en vogels een hersenstamgebied bezitten dat als NAmb wordt aangeduid, missen zij de gemyeliniseerde cardio‑inhibitoire vagale efferenten die bij zoogdieren autonome flexibiliteit en sociale betrokkenheid faciliteren. Bij zoogdieren leidde de ventrale migratie van cardio‑inhibitoire neuronen van de DMNX naar de NAmb tot een gemyeliniseerd cardio‑inhibitoir vagaal subsysteem – een evolutionaire innovatie die adaptieve cardiale regulatie tijdens sociale betrokkenheid mogelijk maakt.
Deze reorganisatie vormt de basis van het VVC – een zoogdierspecifiek subsysteem dat autonome regulatie integreert met motorische banen die het gezicht, het hoofd en de vocale banen aansturen via de craniale zenuwen V, VII, IX, X en XI. Het VVC maakt contextgevoelige cardiale regulatie mogelijk in coördinatie met gelaatsexpressie en vocale prosodie, en levert zo de neurofysiologische basis voor co-regulatie, prosociaal gedrag en relationele veiligheid.
Deze evolutionaire overgang zien we terug tijdens de embryonale ontwikkeling, waarbij cardio-inhiberende neuronen ventraal migreren van de DMNX naar de NAmb. Dit ontogenetische verloop weerspiegelt de fylogenetische transformatie die leidde tot het VVC in zoogdieren, en versterkt zo het idee dat ontwikkelingsprocessen echo’s zijn van evolutionaire aanpassingen.
Cruciaal is dat deze neuroanatomische integratie ten grondslag ligt aan twee kenmerkende eigenschappen van zoogdieren: de coördinatie van zuig–slik–ademhalingspatronen die het zogen mogelijk maken, en het mogelijk maken van vocale prosodie – de modulatie van toonhoogte, ritme en klankkleur die affectieve intenties overbrengt en veiligheid aanduidt. Beide gedragingen vereisen een precieze, toestandsafhankelijke coördinatie van de hersenzenuwen IX (glossopharyngeus), X (vagus) en XI (accessorius), die het strottenhoofd, de keelholte en het zachte gehemelte reguleren. Dit gedeelde circuit weerspiegelt een functionele continuïteit tussen voedingsgedrag in het vroege leven en later opkomende sociale communicatie.
Binnen het polyvagale raamwerk weerspiegelen zowel de mogelijkheid om te zogen als het met prosodische nuances vocaliseren dezelfde onderliggende integratie van hersenstam en viscera die tevens socialiteit ondersteunt. Hoewel de hersenzenuwen V (trigeminus) en VII (facialis) bijdragen aan articulatie, kaakbewegingen en gezichtsuitdrukking, is het de gecoördineerde activiteit van IX, X en XI die zowel de ritmische motorpatronen voor het zogen mogelijk maken, als de dynamische stemmodulatie die co-regulatie en relationele verbondenheid ondersteunt.
Belangrijk is dat zoogdieren en reptielen zich niet langs een lineair continuüm ontwikkelden. Ze splitsten zich af uit een gemeenschappelijke amniote voorouder in afzonderlijke evolutionaire trajecten: zoogdieren uit synapsiden en reptielen uit sauropsiden. Deze vertakking bracht lijnspecifieke innovaties en functionele herbestemmingen van bestaande structuren voort. Pogingen om autonome of sociale functies bij zoogdieren af te leiden uit de anatomie van reptielen gaat voorbij aan de basisprincipes van fylogenetische divergentie. Bij reptielen behoudt de NAmb een somatomotorische rol maar mist zij de integratie met gemyeliniseerde cardio‑inhibitoire vagale controle. Hoewel sommige reptielensoorten beperkte vocalisatie vertonen, beschikken zij niet over de capaciteit voor gecoördineerde, prosodische uitdrukking of voor zuigen die zoogdieren bezitten – gedragingen die afhankelijk zijn van de evolutionair nieuwe koppeling van autonome regulatie met motorische output uit de hersenstam.
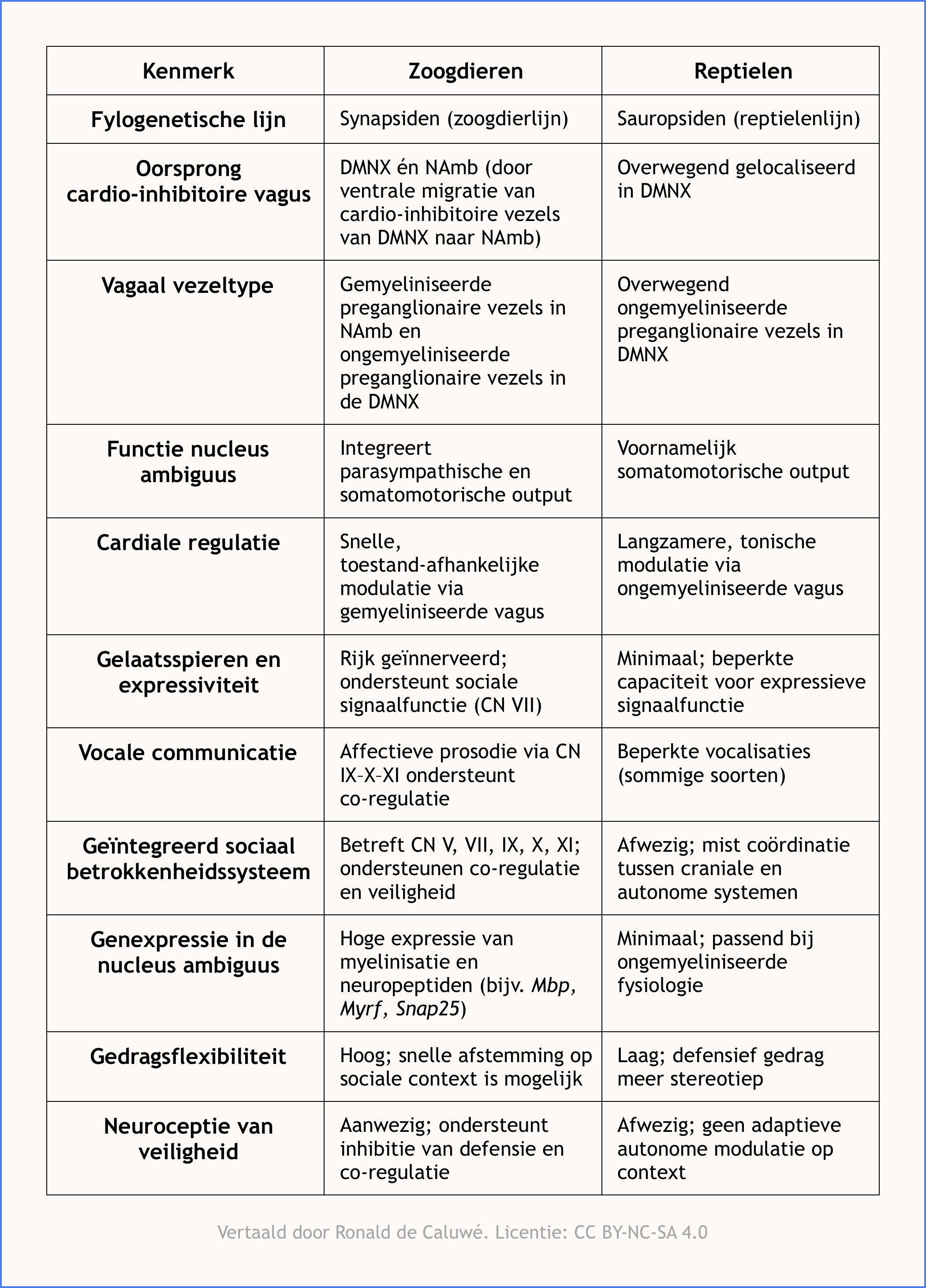
Zoals samengevat in Tabel 1 tonen de neuroanatomische, fysiologische en gedragsverschillen de unieke kenmerken van de autonome en sociale betrokkenheidssystemen bij zoogdieren wanneer we deze vergelijken met die van reptielen.
Tabel 1. Vergelijkende kenmerken van autonome en sociale betrokkenheidssystemen bij zoogdieren en reptielen.
Dit vat de belangrijkste neuroanatomische, genetische en gedragsmatige verschillen samen die relevant zijn voor de polyvagaaltheorie.
Transcriptomische studies versterken deze divergentie. Single‑nucleus RNA‑sequencing bij volwassen muizen laat zien dat neuronen binnen de zoogdier‑NAmb afzonderlijke genprofielen tot expressie brengen die geassocieerd zijn met myelinisatie (bijv. Mbp, Myrf), neuropeptidesignalering en synaptische plasticiteit (bijv. Snap25). Deze expressiepatronen onderscheiden de NAmb van aangrenzende hersenstamkernen en wijzen op haar gespecialiseerde rol in autonome regulatie, vocalisatie en ingestief gedrag (Cao et al., 2019; Liu et al., 2024). Deze moleculaire specialisaties leveren sterk bewijs dat de vocale prosodie bij zoogdieren en de coördinatie van zuigen‑slikken‑ademen niet louter structurele aanpassingen zijn, maar genetisch gebaseerde innovaties in de organisatie van de hersenstam zijn die dynamische sociale betrokkenheid mogelijk maken.
Zoals dit vergelijkende perspectief laat zien, is de autonome organisatie bij zoogdieren op unieke wijze gestructureerd om flexibel, toestandsafhankelijk sociaal contact te ondersteunen – een thema dat verder wordt uitgewerkt in de analyse van evolutionaire innovaties in de volgende paragraaf.
2.3 Vergelijkend bewijs en evolutionaire implicaties
Om de unieke kenmerken van de autonome regulatie bij zoogdieren te begrijpen, is het essentieel de evolutionaire divergentie tussen zoogdieren en reptielen en de implicaties daarvan voor gedrags‑ en fysiologische aanpassingen te beschouwen. Zoogdieren en reptielen hebben zich niet ontwikkeld langs een lineair continuüm; in plaats daarvan zijn zij afgesplitst vanuit een gemeenschappelijke amniote voorouder, wat leidde tot afzonderlijke evolutionaire afstammingslijnen – zoogdieren vanuit de synapsiden en reptielen vanuit de sauropsiden (Pough et al., 2023). Terwijl beide groepen bestaande elementen van het autonome zenuwstelsel behielden, hebben zoogdieren deze structuren aangepast om grotere autonome flexibiliteit in dienst van sociaal gedrag te ondersteunen.
Een cruciale innovatie was het ontstaan van gemyeliniseerde cardio‑inhibitoire vagale efferenten die ontspringen in de NAmb (Porges, 2007; Liu et al., 2024). Deze vezels maakten snelle, toestandsafhankelijke modulatie van visceraal functioneren mogelijk en daarmee dynamische afstemming op omgevings‑ en relationele signalen. Dit markeerde een fundamentele verschuiving van langzame, homeostatische regulatie naar flexibele fysiologische ondersteuning van sociale betrokkenheid, co‑regulatie en emotionele expressie (Porges, 2001, 2007, 2023).
Functioneel ondersteunt deze evolutionaire ontwikkeling gedragingen die zowel biologisch essentieel als sociaal betekenisvol zijn – zoals zogen, vocale prosodie en affectieve communicatie. Deze gedragingen berusten op fijn afgestemde coördinatie tussen de autonome en motorische systemen van de hersenstam, wat illustreert hoe viscerale regulatie en sociale signalen bij zoogdieren nauw met elkaar zijn geïntegreerd (Porges, 2023; Cao et al., 2019).
Het VVC – gedefinieerd door zijn gemyeliniseerde, cardio-inhiberende vagale efferenten – is niet louter een anatomisch kenmerk, maar een cruciale evolutionaire ontwikkeling die het structurele en functionele substraat vormt voor zoogdiersocialiteit. Door snelle, toestandsafhankelijke veranderingen in autonome activiteit mogelijk te maken, stelt het VVC zoogdieren in staat viscerale regulatie te coördineren met gedragsmatige en relationele betrokkenheid. Deze capaciteit om autonoom flexibel te zijn, ligt ten grondslag aan de dynamische regulatie van verbinding, veiligheid en co-regulatie en vormt de neurofysiologische basis van de polyvagaaltheorie, die benadrukt dat de fysiologische toestand onlosmakelijk verbonden is met emotioneel welbevinden en sociaal gedrag (Porges, 2023).
Nu deze evolutionaire verschillen zijn vastgesteld, onderzoekt het volgende onderdeel de neurofysiologische basismechanismen die de autonome integratie bij zoogdieren onderscheiden van die bij niet-zoogdieren, met bijzondere aandacht voor de oorsprong en meting van respiratoire sinusaritmie (RSA).
2.4 Ademfrequentie in RSA, de gemeenschappelijke cardiopulmonale oscillator, en de neurofysiologische grondslagen van de polyvagaaltheorie
De polyvagaaltheorie biedt een fylogenetisch gebaseerd model van autonome regulatie, waarin wordt gesteld dat RSA geen mechanisch artefact van de ademhaling is, maar een neurofysiologische marker van ritmische vagale output die haar oorsprong vindt in de NAmb. De RSA‑frequentie loopt parallel met het ademritme omdat beide worden aangestuurd door een centrale patroongenerator – vaak de ‘gemeenschappelijke cardiopulmonale oscillator’ genoemd – in de hersenstam (Richter & Spyer, 1990).
Deze oscillator organiseert respiratoire en cardiale output via ritmische input vanuit medullaire ademhalingscircuits, met name het pre‑Bötzingercomplex en de nucleus tractus solitarius (NTS), naar zowel de nervus phrenicus (die de inspiratie aandrijft) als cardiale vagale neuronen die de RSA genereren in de NAmb. De n. phrenicus, afkomstig uit cervicale wortels C3–C5, innerveert het diafragma en stuurt de ademhaling synchroon met elke cyclus. Neurofysiologische studies tonen aan dat dezelfde medullaire patroongenerator cardiale vagale efferenten meeneemt, wat resulteert in een strak gekoppelde, fase‑vergrendelde relatie tussen ademhaling en respiratoire ritmen met de hartslag (Richter & Spyer, 1990; Mendelowitz, 1999; Smith et al., 1991; Neff et al., 2003). Deze inzichten bieden de basis om RSA te herframen als centrale – en niet perifere – biomarker van autonome regulatie, een onderwerp dat essentieel is voor de translationele relevantie van de polyvagaaltheorie.
Cardiale vagale neuronen in de NAmb zijn intrinsiek stil tenzij geactiveerd door deze centrale, ademhalingsgekoppelde input, en produceren slechts fasische vagale output als reactie op oscillator‑gedreven excitatie. Zodoende weerspiegelt de RSA‑frequentie het intrinsieke ritme van dit centrale mechanisme – niet perifere feedback of mechanische inflatie, zoals door sommige critici ten onrechte is beweerd (Grossman & Taylor, 2007). Dierstudies bevestigen dat uitbarstingen van activiteit in de n. phrenicus samenvallen met uitbarstingen van cardiale vagale efferente activiteit en ondersteunen zo het model van een gedeelde centrale oscillator.
Dit mechanisme, dat al in de eerste publicaties over de polyvagaaltheorie beschreven is (Porges, 1995; 2007; 2023), identificeert de NAmb als bron van gemyeliniseerde cardio‑inhibitoire output binnen het VVC – een systeem dat uniek is voor zoogdieren en afwezig bij reptielen – en dat bij zoogdieren de RSA produceert. Deze via neuro‑anatomische migratie ontstane vezels geven zoogdieren de capaciteit voor snelle, omkeerbare cardiale modulatie – een voorwaarde voor adaptief, geco‑reguleerd sociaal gedrag en aanpassingsvermogen van gedrag (Porges, 2007, 2023).
Het opnemen van de n. phrenicus als directe effector van de gemeenschappelijke cardiopulmonale oscillator benadrukt de geïntegreerde aard van respiratoire en cardiale controle bij zoogdieren en biedt een neurofysiologische basis voor de dynamische koppeling van fysiologische toestand en sociaal gedrag die de polyvagaaltheorie kenmerkt. Deze centrale coördinatie stelt zoogdieren in staat efficiënt van fysiologische toestand te wisselen in reactie op omgevings‑ en sociale signalen, ter ondersteuning van veerkracht en betrokkenheid op manieren die bij niet‑zoogdierlijke gewervelden niet worden waargenomen.
Deze geïntegreerde, zoogdierspecifieke organisatie van de gemeenschappelijke cardiopulmonale oscillator is fundamenteel voor het begrijpen van RSA als centraal neurofysiologisch signaal. De methodologische implicaties – en hoe dit kan worden gekwantificeerd – worden in de volgende paragraaf behandeld. Deze neurofysiologische inzichten in de oorsprong van RSA creëren een mogelijkheid om het mechanisme om centrale autonome integratie te meten te heroverwegen – een onderwerp dat middels de ontwikkeling van ‘gewogen coherentie’ wordt beschreven.
2.5 Het kwantificeren van centrale autonome integratie: ontwikkeling en herinterpretatie van gewogen coherentie
De ontwikkeling van gewogen coherentie als maat voor centrale autonome integratie (Porges et al., 1980) vormde een belangrijke methodologische vooruitgang in de studie van cardiorespiratoire fysiologie. Met kruisspectrumanalyse beoordeelt gewogen coherentie de fase‑consistentie tussen respiratoire en hartritme‑oscillaties, waarbij deze consistentie wordt gewogen naar de variantie van de hartslag bij elke ademfrequentie. In tegenstelling tot traditionele RSA‑amplitudemetingen (bijv. Lewis et al., 2012) onthult deze dynamische, systeemgerichte index zowel individuele als contextafhankelijke verschillen in autonome functie.
Empirische studies hebben het gebruik van gewogen coherentie als biomarker gevalideerd: hogere coherentie voorspelt grotere anticipatoire hartslagvertraging tijdens taken die aandacht vereisen (Porges & Coles, 1982), en stimulantia moduleren coherentie parallel aan gedragsveranderingen (Porges et al., 1981). Farmacologisch onderzoek toonde verder aan dat, terwijl de RSA‑amplitude daalt bij vagale blokkade, gewogen coherentie stabiel blijft – een afwijking die het best wordt verklaard door een centrale in plaats van perifere oorsprong van de koppeling tussen ademhaling en hartslag (Porges, 1986).
Aanvankelijk werd gewogen coherentie geïnterpreteerd als maat voor parasympathische (vagale) outflow. De empirische afwijking onder cholinerge blokkade leidde echter tot een herinterpretatie. Met verdere uitwerking van de polyvagaaltheorie en de introductie van vagale efficiëntie is duidelijk geworden dat gewogen coherentie, vergelijkbaar met vagale efficiëntie, centrale hersenstamcoördinatie weerspiegelt, aangezien beide in stand blijven onder farmacologische challenge (Porges, 1986, 2024). Gewogen coherentie kwantificeert specifiek de efficiëntie van de centrale cardiopulmonale oscillator, die het pre‑Bötzingercomplex, de nucleus tractus solitarius (NTS) en de NAmb integreert (Smith et al., 1991; Richter & Spyer, 1990, 2001; Mendelowitz, 1999). Deze maatstaf biedt daarmee een directe, functionele beoordeling van de neurofysiologische processen die ten grondslag liggen aan cardiorespiratoire integratie.
Methodologische innovaties zoals gewogen coherentie dagen benaderingen uit die door Grossman en Taylor (2007) worden voorgesteld, die pleiten voor statistische correctie van RSA voor ademhalingsvariabelen, als gevolg van de onjuiste aanname dat RSA een mechanisch artefact van de ademhaling is.
Zoals in de vorige paragraaf uiteengezet, worden RSA en ademhaling co‑gemoduleerd door een centrale oscillator die uniek is voor zoogdieren. Pogingen om ademhaling statistisch uit RSA te verwijderen, verstoren het neurofysiologische signaal en elimineren betekenisvolle variantie die is gekoppeld aan centrale autonome controle. Deze kritieken betichten de polyvagaaltheorie ten onrechte van het ontberen van een neurowetenschappelijke basis en misrepresenteren de centrale rol van de NAmb in het genereren van RSA (Porges, 2023).
Gewogen coherentie biedt daarmee een robuuste maat van centrale autonome integratie op systeemniveau, en verduidelijkt zo belangrijke neurofysiologische en evolutionaire vraagstukken (Porges & Bohrer, 1980; Richter & Spyer, 1990). De bredere implicaties van deze innovaties voor zowel de vergelijkende biologie als de klinische wetenschap worden besproken in de volgende paragraaf.
2.6 Evolutionaire, klinische en translationele implicaties van methodologische innovatie
De validatie van centrale indexmaten zoals gewogen coherentie en vagale efficiëntie (Porges, 1986, 2023, 2024) heeft diepgaande implicaties voor zowel de evolutionaire biologie als de klinische wetenschap. Door een directe index van centrale autonome regulatie te bieden, verduidelijken zij de oorsprong en functionele betekenis van RSA, en bevorderen ze daarnaast het begrip van zoogdierspecifieke aanpassingen in autonome controle.
Bij zoogdieren maken gemyeliniseerde cardio‑inhibitoire vezels binnen het VVC – ritmisch ‘gepoort’ door medullaire ademhalingscircuits – snelle, contextgevoelige modulatie van cardiale output mogelijk (Porges, 1995, 2007, 2023; Mendelowitz, 1999; Neff et al., 2003). Dit onderscheidt zoogdieren van reptielen, waar cardiale vagale controle primair wordt gemedieerd door ongemyeliniseerde vezels vanuit de DMNX, en waar een gecoördineerde cardiopulmonale oscillator ontbreekt (Richter & Spyer, 1990; Porges, 2023). Dientengevolge zijn bij deze soorten interacties tussen ademhaling en hartslag minimaal of afwezig als marker van centrale cardiale sturing. Hun autonome regulatie is vooral gekoppeld aan het voldoen aan de metabole eisen van beweging en blijft grotendeels losgekoppeld van sociale interactie (Porges, 2007, 2021; Liu et al., 2024).
De translationele waarde van centrale autonome indexwaarden blijkt uit hun toepassing als betrouwbare, niet‑invasieve biomarkers voor autonome functie in uiteenlopende klinische contexten, waaronder trauma, angst, autisme en andere neuro‑ontwikkelings‑ of psychiatrische aandoeningen (Porges et al., 1981; Porges, 1976, 2023, 2024). Deze indices zijn bruikbaar in verschillende settings, en hun robuustheid onder farmacologische uitdaging benadrukt hun onderzoeks‑ en klinische waarde (Porges, 1986, 2024).
Ondanks de oproepen van Grossman tot statistische correctie van RSA voor ademhaling (bijv. Grossman & Taylor, 2007), gaan hun argumenten voorbij aan zowel het evolutionair unieke karakter als aan het centrale, door de hersenstam gemedieerde mechanisme van RSA (Richter & Spyer, 1990, 2001; Porges, 2023, 2024). Door RSA te herdefiniëren vanuit het perspectief van de polyvagaaltheorie en gebruik te maken van gevalideerde centrale maatstaven, wordt een samenhangende basis gelegd voor toekomstig translationeel onderzoek, klinische diagnostiek en therapeutische toepassingen. Door de focus te leggen op systeemniveau-indicatoren zoals gewogen coherentie, bevordert de polyvagaaltheorie zowel het evolutionaire inzicht als de ontwikkeling van effectievere interventies voor aandoeningen die geworteld zijn in verstoorde autonome regulatie.
Einde van het eerste deel van drie.
Download het oorspronkelijke artikel hieronder.
Als je geïnteresseerd bent in de polyvagaaltheorie, volg dan ook het Polyvagaalinstituut Nederland.
Als je dit artikel lezenswaardig vond en (nog) geen betaald abonnement wilt, mag je me ook trakteren op een cappuccino!
Ja, af en toe had ik er spijt van …
En dan is er nog een literatuurlijst, maar die laat ik logischerwijs onvertaald.
Dit betekent dat iedereen mijn vertaling mag overnemen, alleen voor niet-commerciële doeleinden, met vermelding van de auteurs- en vertalersnaam. Ziehier voor de volledige rechten en plichten.
Het betekent ook dat deze artikelenserie dus niet achter de betaalmuur terecht gaat komen.
Referentie: Porges, S. W. (2025). Polyvagal Theory: Current Status, Clinical Applications, and Future Directions. Clinical Neuropsychiatry, 22(3), 169–184.
DOI: doi.org/10.36131/cnfioritieditore20250301
Wat dapper om zijn stukken te vertalen! Porges staat er om bekend dat zijn uitleg stevige kost is. Ook mondeling. Niet voor niets is de vertaalslag zoals Deb Dana het doet zo aansprekend.
Ik heb het gelezen maar om het te laten landen moet ik het nog een keer lezen. In elk geval is het voor mijn in het Nederlands beter te begrijpen dan in het Engels.
Dus bedankt voor je inspanningen!
Bedankt, ik zie uit naar de de vertaalslag van deze 3 artikelen die je wilt gaan schrijven.