

How Old Is That Ventral Vagus, Really?
Older than Porges suspected, smarter than Taylor names. Unraveling a scientific debate about polyvagal theory


Lees je liever de Nederlandse versie?
Welcome to my longest article on RelaxMore.net so far. It cost me a fair amount of work, but it was worth it. Working through the scientific sources was intensive, instructive, and enjoyable. But most of the work went into finding the wording that does justice to both sides of a debate that has been running for almost thirty years and that I am now writing about. Take your time to read it. Because it is a long piece, I will begin with a summary. I also hope it will leave you all the more curious about how the story unfolds.
Summary
Since the mid-1990s, there has been debate about the evolutionary foundation of polyvagal theory. On one side stands Stephen Porges, who presents the ventral vagal system (= the social engagement system) as typically mammalian: a new, fast vagal brake on the heart, originating in the nucleus ambiguus, that enables mammals to seek social contact and to calm one another. On the other side stands a group of comparative physiologists led by Edwin Taylor, who have shown that the building blocks Porges names—myelinated cardiac vagal fibers, two nuclei in the brainstem, and a respiratory-modulated heart rhythm (I explain all the terms further on)—already existed hundreds of millions of years before mammals, in sharks, lungfishes, frogs, and reptiles.
At first glance this looks like a discussion that has to produce a winner. But the opposition turns out to be far less spectacular once you apply the evolutionary concept of exaptation: the phenomenon whereby an existing structure acquires a new function in a new context. The building blocks are therefore old, but what mammals have done with them is indeed new: an integration of heart, breathing, and voice into one coherent regulatory system, organized around something no other vertebrate possesses to this degree, a mobile and expressive face.
In this article I show how that perspective unites Porges and his critics within a single evolutionary framework and how it makes polyvagal theory, paradoxically, more empirically testable than it was in its original form.
A thirty-year disagreement
Since the mid-1990s, there has been debate about the evolutionary foundation of polyvagal theory (Porges, 1995, 2011). On one side stands Stephen Porges, the American scientist who developed the theory. He argues that mammals, and therefore we humans too, possess a special neural pathway that other animals do not have.
This neural pathway1 is part of the tenth cranial nerve, the well-known vagus nerve. According to Porges, this pathway makes it possible for mammals to find rest in one another’s company and to convey safety to one another through their body language, voice, and facial expression. It is the neural, bodily, and behavioral foundation of what he calls the social engagement system.
On the other side stands a group of comparative physiologists, researchers who compare the nervous systems of different animal groups with one another. The British emeritus professor Edwin Taylor (University of Birmingham), together with Tobias Wang (Aarhus University) and Cleo Leite (Federal University of São Carlos), has for decades carried out research into the regulation of heartbeat and breathing and the coordination thereof in fishes, frogs, lizards, and birds2. In 2022 they published an article titled “An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the ‘Polyvagal Theory’” (phylogeny being the study of the evolutionary descent of a group of organisms, and cardiorespiratory control the regulation of heartbeat and breathing).
Their objection to polyvagal theory is not that Porges’s functional account of social behavior is nonsense. Their objection runs deeper: they say that the building blocks on which Porges bases the system—contrary to what he claims—do not occur exclusively in mammals at all. Those nerve fibers, those brainstem nuclei, that interplay between breathing and heartbeat: you already find these in fishes, lizards, and frogs as well. Taylor, Wang, and Leite describe this position at length in their, incidentally very interesting, article (Taylor, Wang, & Leite, 2022; see above for a PDF). Earlier, Paul Grossman had already advanced a methodological critique pointing in a similar direction (Grossman & Taylor, 2007; Grossman, 2023).
At first glance this looks like a fight that can have only one winner. Either the building blocks are typical of mammals, or they are not. But anyone who conducts the debate in this either-or fashion misses something important. There is a third possibility that does justice to both sides, a both-and model. The room for it lies hidden in an evolutionary concept called exaptation, a word I explain further on. With the help of that concept, the opposition turns out to be less irreconcilable than it seems.

We begin with the basics
Before we dive into the debate itself, I will first explain a few concepts so you can follow the reasoning.
The vagus nerve
The vagus nerve is the tenth cranial nerve. It is a long, fairly thick nerve that runs down from the brainstem and branches out to the heart, the lungs, the larynx, and the organs in the abdomen. It is the longest nerve in the body. The vagus belongs to the parasympathetic nervous system: the part of our nervous system that is active in rest and recovery, that slows the heartbeat, supports digestion, and is involved in voice and facial expression, but also in shutdown and collapse responses under stress that we experience as life-threatening.
The vagus does not have one origin in the brainstem, but two. In the brainstem there is a cluster of nerve cells situated more towards the back: the dorsal (= back side) motor nucleus of the vagus, abbreviated DMNX. And there is a cluster of cells situated more towards the belly side (= ventral): the nucleus ambiguus, abbreviated NAmb. That these are two different brainstem nuclei is important in polyvagal theory. Porges links them to different functions. The DMNX is, in his view, evolutionarily old and is associated with collapsing, shutting down, and dissociation under overwhelming threat, which in polyvagal theory is called shutdown or collapse. The NAmb, by contrast, is said to have a function in mammals that no other animal group possesses. Not so much because this nucleus would be absent elsewhere (Porges himself acknowledges that an NAmb-like structure is already present in reptiles), but because the fast, myelinated fibers (the next concept I will explain) that run from the NAmb to the heart, and the way they are connected to face, voice, and hearing, are, in his view, unique to mammals. This NAmb branch is said to make it possible to seek social contact, to calm down, and to be in connection.
A word of warning is in order here, because I just used the word “branch,” and that word is actually misleading. You encounter it everywhere in the literature on polyvagal theory: the ventral vagal branch, and the dorsal vagal branch, as if these were two separate cables each finding its own way through the body. That is not how it works. The fibers that come from the DMNX and the fibers that come from the NAmb do not run separately but become interwoven into the same nerve strand even before leaving the brainstem. So do not picture two loose wires, but one rope in which threads of different origin are braided together. The cardiac branch of the vagus, which runs to the heart, therefore contains fibers from both nuclei, fast myelinated and slow unmyelinated, mixed together (Taylor et al., 2022). Incidentally, I will keep speaking of “branches” in this article because it is a handy and well-established word, but you now know it is a simplification.
That the two types of fibers can be distinguished functionally therefore does not mean that they are anatomically separate. The distinction lies in their origin and in their behavior, not in two loose branches. This may seem a detail, but it touches on something that recurs throughout this article: a functional distinction is something other than a physical separation. Anyone who imagines the ventral and the dorsal vagus as two separate neural pathways makes exactly the thinking error that also underlies part of the criticism of polyvagal theory. I will return to this further on.
Myelin
One more concept is important: myelin. Some nerve fibers are wrapped in an insulating layer of fat and protein. That layer, myelin, allows signals to travel much faster through the fibers. Compare it with the plastic insulation around an electrical wire. Nerve fibers with myelin conduct up to ten times faster than fibers without myelin. An important claim of Porges about the control of the heart is that only mammals have myelinated NAmb fibers for this purpose, and that these fast fibers form the basis for the delicate interplay between breathing and heartbeat that expresses itself in heart rate variability.
That interplay itself is called respiratory sinus arrhythmia, abbreviated RSA. It is the natural phenomenon whereby the heart speeds up a little on inhalation and slows down a little on exhalation. It is often considered a sign of a healthy, flexible nervous system.
What Taylor and colleagues have shown
With these concepts in hand, we can look at what Taylor, Wang, and Leite published in 2022 in the journal Biological Psychology (Taylor et al., 2022). Their review article condenses decades of research into a structured critique of the evolutionary underpinning of polyvagal theory. Their argument has four main lines.
The first line concerns the inhibition of the heart at rest. In all vertebrate animals studied, from sharks to mammals, the heartbeat at rest turns out to be slowed by the vagus. In reptiles this brake even turns out to be very strong. In the rattlesnake the heart rate roughly doubles when the vagal influence is removed with atropine (Campbell et al., 2006), whereas in humans the increase usually stays between thirty and fifty percent. A slow metabolism, therefore, goes perfectly well together with a powerful vagal grip on the heart. That takes some getting used to for the intuition, which expects a cold-blooded, slow animal to have a loose rather than a tight rein. Strong vagal control of the heart at rest is, in any case, no mammalian novelty. Whether that control in reptiles also rests on the same fast, myelinated fibers as in us is a separate question, to which I return at the third line.
The second line concerns the two origins of the vagus in the brainstem. Porges presents it as a typically mammalian feature that vagus fibers controlling the heart arise from two nuclei in the brainstem: in the DMNX and in the NAmb. But Taylor and his colleagues show that this division already exists in the dogfish. In this shark the greater part of the vagal nerve cells lies in the DMNX, but there is also a cluster situated more towards the front (= ventral), and in that cluster there already lie cells that control the heart. Taylor and colleagues see this ventrolateral group as a possible precursor of what in mammals has become the nucleus ambiguus. In crocodiles, lizards, snakes, and frogs, the authors find comparable distributions. In the African clawed frog, as much as 30% of the relevant vagal nerve cells already lie in that front brainstem nucleus. The division is not only old; in the ventral group of the shark there already lie cardiac vagal neurons, at a location that Taylor regards as a possible precursor of the mammalian NAmb. And its relative size increases through evolution, from ten percent in the shark to thirty in the clawed frog and an extensive nucleus in us. So not an invention of “ours,” but an expansion.
The third line concerns the myelinated fibers. Porges’s claim here is specific: not that myelin would be present somewhere in the vagus, but that only mammals possess fast, myelinated cardiac fibers that can fine-tune the heart from moment to moment within a respiratory-modulated (= driven by breathing) circuit. Literally: “Only mammals have a myelinated vagus” (Porges, 2009, 2011). With the electron microscope, Taylor and colleagues have meanwhile demonstrated these fibers in the cardiac vagal branch in sharks, lungfishes, rattlesnakes, lizards, and toads as well. In the dogfish and the South American lungfish, the conduction velocities, moreover, turn out to match those of certain fibers in mammals. And in both the lungfish and the rattlesnake, these fibers take part in a respiratory-modulated heart rhythm that resembles the RSA we know in mammals. So this is not a matter of a few exceptions where myelin has been found “somewhere,” but of the same functional package that Porges presents as mammalian.
The fourth and theoretically most interesting line concerns the interplay between breathing and heartbeat itself. Taylor and his colleagues call this cardiorespiratory synchronization (CRS): each heartbeat coincides with a breathing movement, a one-to-one coupling, controlled from the DMNX3, which optimizes the counter-flow of blood and water across the gills for maximum oxygen uptake. In air-breathers, such as lungfishes, frogs, reptiles, birds, and mammals, the coupling between breathing and heartbeat looks fundamentally different. The heartbeat is then much faster than the breathing rhythm, so a strict one-to-one coupling is no longer possible. Instead, breathing influences the speed of the heartbeat: during inhalation the vagal brake on the heart is briefly released so that the heartbeat accelerates, and during exhalation the brake is applied again. This pattern, in which the heart rate fluctuates to the rhythm of breathing, is what in mammals we call respiratory sinus arrhythmia (RSA). This same mechanism has thus been found in lungfishes and cururu toads, animals whose lineage split off hundreds of millions of years ago from the line that would eventually lead to mammals4.

The most surprising insight lies in the authors’ concluding thought. In reptiles and amphibians, which have a heart in which the pulmonary and systemic circulations are not fully separated—as is the case in us—this fluctuation in heart rate with each breath helps to send blood efficiently to the lungs5. Mammals no longer need that efficiency gain: our heart has fully separated atria and ventricles for the pulmonary circulation and for the systemic circulation. RSA in mammals is therefore actually not an evolutionary innovation. It is a remnant from a time when the rhythm still had real work to do and still beats today 😉, but without the clear physiological role it still fulfills in animals with cardiac shunting.
Here a question may arise. If RSA in mammals has lost its original function, why then does heart rate variability (of which RSA is an important component) count as a measure of health? The answer is that RSA in us is valuable not for what it does, but for what it reveals. The fluctuation of the heartbeat with the rhythm of breathing arises because the fast ventral vagal fibers continually fine-tune the heart. The magnitude of that fluctuation is thereby a window onto how smoothly that vagal regulation works. A lot of variability points to a flexible, responsive system; little variability to a system that falters through stress, illness, or age. RSA in mammals is therefore no longer a functional mechanism but a usable and measurable signal, and in that capacity it is valuable.
What Taylor explicitly does not claim
In the article by Taylor and colleagues, there are a few remarks that often remain underexposed. In their introduction and in their conclusion, the authors say explicitly that they do not wish to say anything about the role of the vagus in human social behavior. They literally write that they do not claim to be able to “say anything useful about the possible roles of a social engagement system in humans that might in some way involve parasympathetic input via the vagus nerve” (Taylor et al., 2022). At the same time, they leave no doubt that they regard the “smart vagus” concept itself, with which Porges grounds that social engagement system, as superfluous. Their delimitation therefore concerns only the psychological and clinical elaboration of the theory; about the neurobiological building blocks beneath it, they do speak out. They acknowledge that the vagus fulfills many important functions in us. Their criticism is confined to what they have themselves studied: the comparative physiology of the heart and breathing in different animal groups.
This is an essential delimitation of their field of research and scientifically very sound. What Taylor and his colleagues refute is the claim that the anatomical building blocks are typical of mammals. What they therefore do not contradict is the claim that mammals have an integrated system in which face, voice, and heart work together to make social contact possible. Here it already becomes apparent that polyvagal theory and the criticism of Taylor and his colleagues need not exclude one another.
Lungfish?
At this point, as a level-headed reader, you may wonder, what does a lungfish have to do with me? My ventral vagus regulates how I deal with colleagues, my capacity to calm my child, and my voice in a conversation. A lungfish does not flirt, does not smile, and does not comfort a partner. What is it doing here in the story, then?
The answer lies in a distinction that matters in evolutionary biology. When a trait occurs in different lineages, it can have arisen in two fundamentally different ways. It can have been invented independently, each time anew, in each lineage separately. That is called convergent evolution: bats, birds, and pterosaurs all three have wings, but each along its own evolutionary path. The common ancestor had no wings. Or it can have been inherited from a common ancestor that already possessed the trait, after which all lineages retained it. That is called homology.
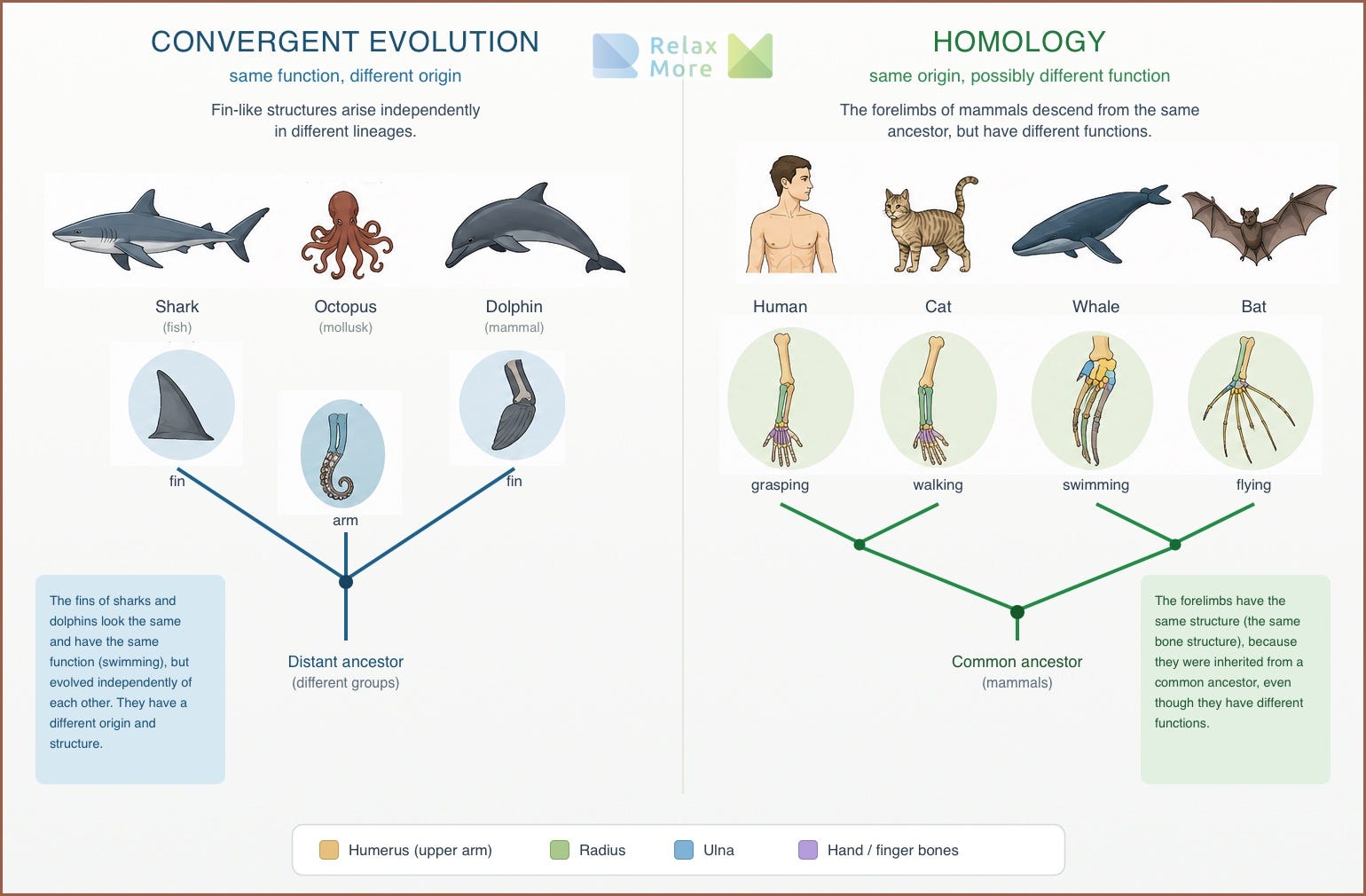
As you can see above, the difference between convergent and homologous seems easy to delineate, but in practice the two do sometimes blur into each other. Take the eye. A light-sensitive organ is estimated to have arisen independently dozens of times in the animal kingdom, from the compound eye of insects to the camera eye of vertebrates and squids. At the level of construction, the eye is therefore a textbook example of convergence: invented again and again, with different parts and different optics. And yet there is a surprise underneath. All those independently arisen eyes turn out to be switched on by the same genetic switching mechanism: the so-called Pax6 gene plays a key role in the formation of eyes in virtually all animals. That gene is homologous, inherited from a distant common ancestor6.
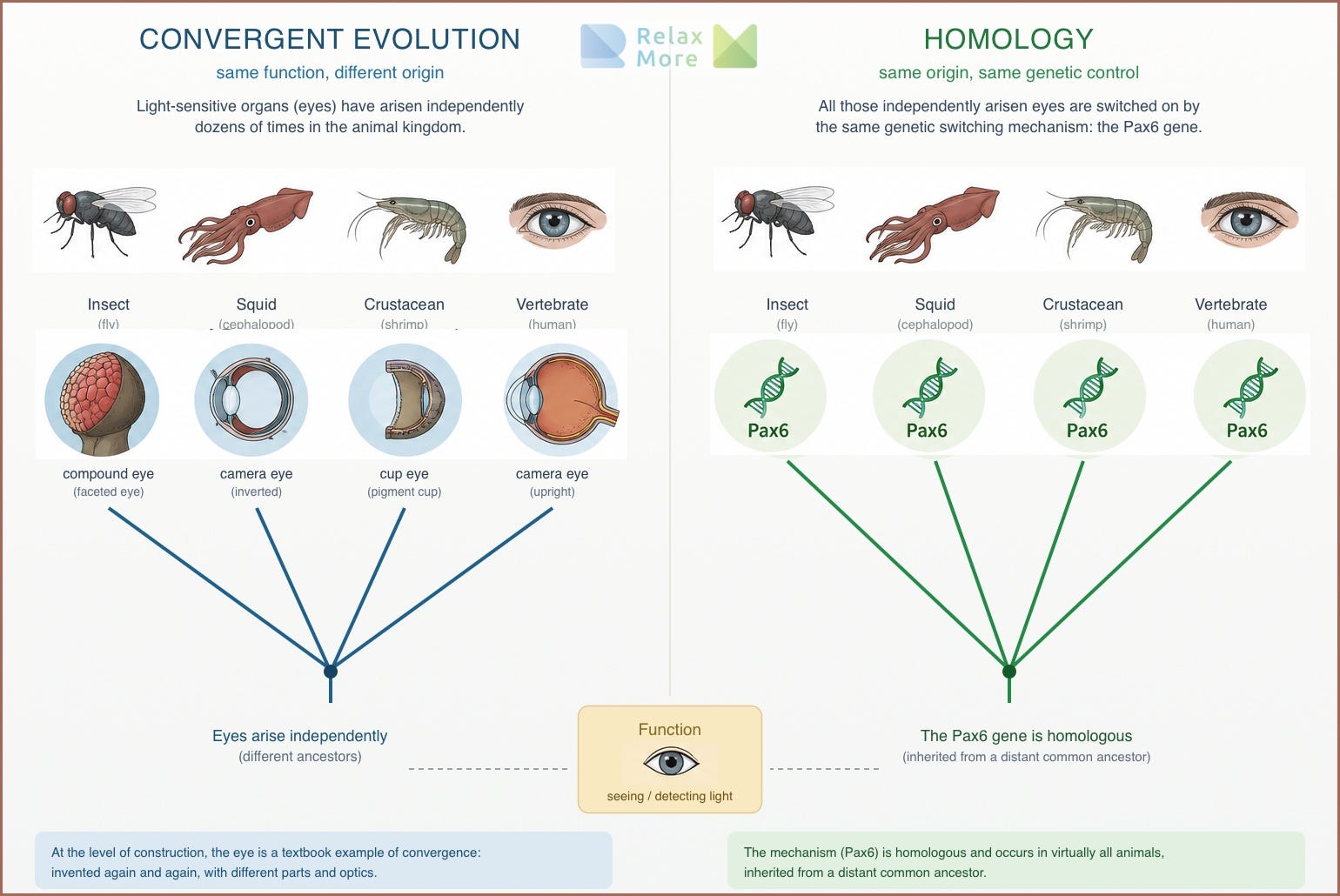
The eye is therefore at once convergent and homologous: convergent in its construction, homologous in its genetic control. The ancient gene gave, in different animals, the starting signal “build an eye here,” and each lineage filled in that command with its own means. Precisely this difference between levels is what we need to understand the vagal story. Because there, too, it holds: you must know at which level you are looking before you can say whether something is old or new.
Taylor and his colleagues take a very clear position: the mechanism of the vagal brake on the heart, with myelinated fibers and respiratory modulation, is homologous. It was not invented independently by lungfishes, toads, and mammals but inherited from a common ancestor that lived somewhere in the Devonian, about four hundred million years ago. That ancestor already had the mechanism: the fast fibers, the cells that control the heart, and the coupling with the breathing rhythm. And all its descendants, each via its own evolutionary trajectory, received it and passed it on in slightly modified form.
This is a more important point than you might say at first glance. If the trait had arisen convergently, Porges could still maintain that mammals developed it anew, as a unique invention on their own evolutionary path. But homology means that it is ancient heritage. The anatomical building blocks, a ventral group of cardiac vagal neurons with myelinated fibers to the heart, are not typically mammalian. They are already present in recognizable form in the dogfish and recur in virtually all later vertebrates. And the RSA-like coupling of heart rhythm and breathing that mammals show, they share with lungfishes, frogs, reptiles, and birds. The mechanism is therefore not new in mammals; it is ancient.
And here, then, it does indeed touch the human being. Not because we have anything to do with the lungfish in our daily life, but because it undermines the timeline of Porges’s theory. Porges bases his theory on the claim that mammals developed a new building block that makes the social engagement system possible. If that building block was already present in the lungfish, that account is no longer tenable in its original form.
It then becomes time to reconsider what mammals really do that is new.
And that is precisely what I am working towards with this whole article: not the building blocks, but what mammals have done with them, is new. The lungfish therefore belongs in this story as a witness to an ancient inheritance that we share with it. The question then becomes: how can we understand what mammals have done with that inheritance?
Here an evolutionary concept comes into play that is called exaptation.

Exaptation
In 1982 the American evolutionary biologist Stephen Jay Gould and his colleague Elisabeth Vrba published a short but influential article under the title “Exaptation, a missing term in the science of form” (Gould & Vrba, 1982). Their thesis was that evolutionary biology had, until then, been missing an important category of form and function.


Until then, one spoke mainly of adaptations: traits that arose through natural selection for the sake of the function they fulfill. The long legs of a horse are an adaptation for running fast; the webbed feet of a duck are an adaptation for swimming. But Gould and Vrba pointed out that many traits that now fulfill a particular function did not arise for the sake of that function. They were shaped in a different environment, for a different purpose, and only later came to fulfill a new function.
For this type of form and function, they coined the term exaptation7.
The term “exaptation” is formed by analogy with “adaptation.” Where the prefix ad- (= towards) points to a trait that has been shaped towards a purpose, ex- (= out of) points to a trait that comes from somewhere else and only later turned out to be useful.
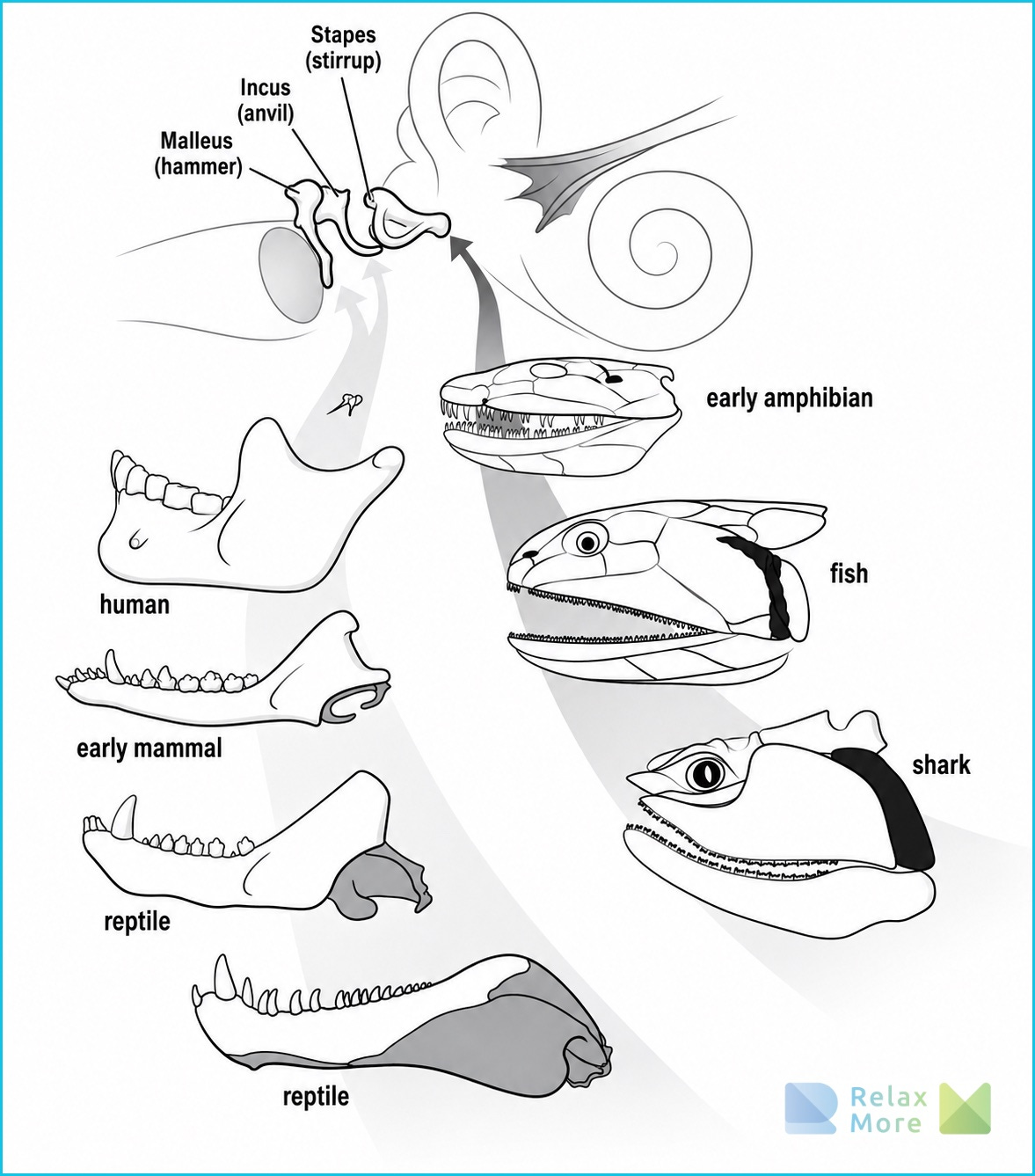
A few examples are probably clarifying: bird feathers almost certainly did not arise for flight. They arose in feathered dinosaurs, presumably for heat insulation or for the sake of sexual display. Only much later, when some species climbed into the treetops and began to make gliding flights from there, did these feathers acquire a repurposing for flight. The middle-ear bones of mammals (the hammer, the anvil, and the stirrup that pass sound on to the inner ear) once arose as bones in the jaw of early precursors of mammals. When the jaw construction changed, those bones lost their old function and were repurposed in hearing. Lungs probably evolved from gas-exchange organs in early fishes that lived in oxygen-poor water; only later were they used for breathing on dry land.
Purposeless, yet reconstructable
The core of exaptation is simple: a trait is older than the function it now fulfills8; it was already there before it became useful. With an adaptation, by contrast, the trait spread precisely because it was useful: ducks with better webbed feet turned out to have greater chances of survival, so webbed feet became more common over the course of many generations. With an exaptation, the trait already existed for another reason, or without any particular reason, and only later turned out to be useful for what it now does. Think again of the feathers of birds, which arose in dinosaurs that could not fly and only much later turned out to be usable for an entirely new function: flying. The feather, then, was there before flight.
With neither adaptation nor exaptation, there is any plan or intention; evolution, after all, strives towards nothing. The distinction is therefore not about goal-directed versus purposeless, but about the question: was a trait already useful while it was arising, or did it exist before it became useful? And how do you establish that if no one was there to witness the origin?
The answer is that you reconstruct it from evidence. Three kinds of clues help here. The first is the order in time: if a trait already existed in ancestors before it acquired its current function, then it is older than that function. Think again here of the feathers in feathered dinosaurs that could not fly. They prove that feathers were there before flight, and feathers are therefore an exaptation. The second clue is the form itself: a trait that developed together with its function is usually well attuned to it, whereas a repurposed trait often bears traces of its old function9, or one whose old function even remains a riddle. The third clue is the comparison between species: by looking at which relatives do and do not have the trait, you can reconstruct when and in what form it originally arose.
The distinction between adaptation and exaptation is therefore not a matter of guessing at intentions but an empirical10 question to be answered via fossils, family trees, and comparative research. And it is precisely this that also makes it testable.
And then there is “integration”
One concept is still indispensable here because it highlights a side that must not be missed in the whole discussion: integration. This is a functional term that points to the phenomenon whereby separate parts together do something they cannot do separately11. Heart, breathing, voice, face, hearing, and social orientation have each been studied separately by different disciplines. But in mammals they work as one coherent regulatory system. This difference in level of analysis is precisely what makes Porges and the comparative physiologists talk past one another.
Taylor and his colleagues study separate parts: nerve fibers, brainstem nuclei, conduction velocities. Porges looks at what those parts produce together. Both approaches are important and legitimate; they simply operate at different levels, and when they enter into debate with one another as if it concerned the same level, misunderstandings easily arise.
One of those misunderstandings I announced earlier, with the word “branch.” Anyone who imagines the ventral and the dorsal vagus as two separate neural pathways wrongly translates a functional distinction into a physical separation. And precisely such a confusion of levels, one moment talking about separate fibers, the next about a coherent system, feeds part of the opposition between Porges and his critics. Much of what is presented as incompatible turns out, on closer inspection, to be a matter of talking at different levels.
Porges’s own reformulation: a half step towards exaptation
An interesting development in this debate is that Porges himself, since about 2021, has in his formulations come very close to the exaptation idea. Without using the term, he does move conceptually in that direction.
In his 2021 article in the journal Comprehensive Psychoneuroendocrinology, Porges no longer speaks of a “unique mammalian system” but of a “repurposed” system (Porges, 2021). Strictly speaking, an intention still resonates in that word, as if something is deliberately given a new task, whereas we know that exaptation takes place without a plan. But apart from that nuance, the move Porges makes is remarkably close to the exaptation idea.
Examples of his formulations: “The nervous system was repurposed to suppress defense strategies in favor of social behavior.” Elsewhere he writes that evolution is said to have repurposed the mammalian vagal complex to facilitate social behavior. In his 2022 article in Frontiers in Integrative Neuroscience, he says it even more explicitly: “Evolution is a process of modification in which existing structures and circuits are adapted to fulfill adaptive functions.” (Porges, 2022). That last is almost word for word what Gould and Vrba meant in 1982 with exaptation. With it, Porges in fact acknowledges that the building-block level is untenable and shifts to the synthesis level. But he does not label this as a correction of his earlier claims.
A second shift concerns the anatomical reference point. In his original work from the 1990s, Porges spoke of the NAmb, the brainstem nucleus that lies somewhat more towards the front (that is, ventral) and from which the faster vagal fibers arise, as the source of what he called the “smart vagus” (Porges, 1995, 2011). In his work from 2021 onwards, he speaks consistently of the ventral vagal complex, a larger group of nerve cells that includes not only the NAmb but also the brainstem nuclei that control the facial muscles, the jaw muscles, and the little muscles in the middle ear (Porges, 2021, 2023).
What is specifically mammalian, therefore, lies not in one separate brain nucleus, nor in a larger collection of nuclei as such. It lies in something that makes mammals unique among the vertebrates: a mobile, expressive face, and in the way old vagus building blocks have come to work together with it. Most mammals have real facial musculature, the muscles under the skin with which we smile, frown, and let our gaze speak. Fishes, reptiles, and birds lack these entirely; a lizard cannot frown, a bird cannot contort its face12. That new face calls for control, and it is that control that in mammals became interwoven with the old regulation of the heart.
At the same time, Porges leaves standing other statements that this half step has not resolved. He continues to write about “the transition from asocial reptiles to social mammals” (Porges, 2021). That is not entirely correct empirically. Researchers such as Doody, Burghardt, and Dinets devoted a whole book in 2021 to the social life of reptiles, with parental care, group formation, and collective nest protection (Doody, Dinets, & Burghardt, 2021). In 2023 they published an article in Biological Psychology in which they directly refute the assumption that reptiles are asocial (Doody, Burghardt, & Dinets, 2023). The exaptation formulation I propose here resolves this tension: it lets social behavior in reptiles stand in its own right, without thereby denying the specifically mammalian interplay of face, voice, and heart.
A final issue concerns the scientific status of the theory itself. In 2021 Porges wrote, “The theory was not intended to be proven or refuted, but to be informed by research and modified.” (Porges, 2021). That is, in terms of the philosophy of science, a problematic statement, one that critics have also regularly held against Porges. A theory that withdraws from testing (which, incidentally, is not what Porges said nor meant) places itself outside science13. Here the exaptation form of polyvagal theory offers an unexpected way out. It actually makes the theory more testable rather than less. The central claim shifts towards the presence and operation of a coherent face-voice-heart system in mammals, and away from anatomical statements that comparative physiology has by now refuted. What remains is open to empirical investigation14.
Polyvagal theory as a theory of exaptation
When we lay the Taylor article and polyvagal theory side by side and view them through the lens of exaptation, the apparent opposition dissolves into a workable synthesis.
It is clarifying here to see that exaptation works on three levels at once and that it is meaningful to keep those levels apart.
At the evolutionary level, exaptation is a form-and-function interpretation: old building blocks acquire a new role in a new environment. At the neurobiological level we see a reorganization: existing networks that become coupled differently acquire a different hierarchy and come to communicate with one another in new ways. At the functional level we see the integration of heart, breathing, voice, face, and hearing that come to operate as one coherent system.
These three levels do not describe three different phenomena, but one phenomenon from three perspectives. Anyone who looks only at the level of the separate building blocks (as sometimes happens in the discussion) sees the continuity with other vertebrates and concludes that nothing special is going on in mammals. Anyone who looks only at the functional level sees the special character but misses the evolutionary continuity. Only when the three levels remain in view at the same time does the story fall into place.
The building blocks that Porges initially presented as typically mammalian (myelinated vagus fibers, a ventral group of heart-controlling vagus cells, a breathing rhythm that modulates the heartbeat) are, empirically speaking, not inventions of mammals. They form part of a much older equipment that all vertebrates share. But what is indeed new in mammals is the way these old building blocks have been integrated into a coherent ensemble that exists nowhere else in the animal kingdom in this form.
In mammals
What is really new in mammals is therefore not the separate building blocks, but the combination. And this combination shows itself most clearly in something no other vertebrate animal possesses: a mobile, expressive face. That face itself is indeed a new anatomical acquisition; what is called exaptation lies not in that face as such, but in the way old vagus building blocks have come to work together with it.
Mammals are the only animal group with real facial musculature, the muscles under the skin with which we smile, frown, blink, and let our gaze speak. These muscles in mammals are diverse and differentiated and are connected with the movements of lips, jaw, tongue, and larynx. Fishes, reptiles, and birds lack this entirely: a lizard cannot frown, and a bird cannot contort its face. On top of that, the larynx develops into a refined apparatus for voice and intonation, and the middle ear acquires a control system that can dampen or amplify sound frequencies, attuned to the sounds that conspecifics make. In babies, several cranial nerves coordinate sucking, swallowing, and breathing together, a pattern that exists in this form in no reptile or amphibian.
That all of this became possible in mammals is connected with a second characteristic: warm-bloodedness. This is, incidentally, not unique to mammals. Birds are also warm-blooded, and there are strong indications that the common ancestors of mammals, birds, and crocodilians possessed a form of warm-bloodedness (Grigg et al., 2021). A warm-blooded animal maintains its body temperature. It can thereby permit itself something a cold-blooded animal cannot, namely a continuous, delicate fine-tuning of the physiological state, partly dependent on what is happening socially. That permanent fine-tuning costs energy, and it is precisely that energy that a warm-blooded animal has available. Warm-bloodedness is thereby not itself the innovation, but a precondition that makes the innovation possible.
And here it becomes visible what the exaptation precisely is. Not the facial expression itself, for that is a new acquisition. And not the vagal brake on the heart; that is, in fact, ancient, and we share it with lungfishes and reptiles. The exaptation is the connection between those two: an old mechanism, the fast vagal brake that can fine-tune the heart from moment to moment, that comes into the service of something utterly new, namely a face that can show an inner state and read it in another. The old heart regulation acquired a social role that became possible only once there was an expressive face to communicate with and once there was enough energy to let that attunement take place continually. This is exaptation in its clearest form: an old part that, in a new configuration, acquires a new meaning.
And in fact that repurposing reaches further than this single coupling of heart and face. For the social engagement system is built up out of a whole series of old parts. The middle-ear bones with which we make out another’s voice arose, as we saw earlier, as fragments of the jawbones of reptiles and sharks and only later acquired their role in hearing. The larynx, with which we shape vocal timbre and intonation, goes back to a valve that closes off the lower airway, a function it still fulfills; the formation of voice came only later. And the vagal brake on the heart is hundreds of millions of years old. Some of these parts, moreover, were already connected with one another earlier. In diving reptiles, the larynx and heartbeat work together to save oxygen during a dive. The separate pieces, and even some of their connections, are therefore not an invention of mammals.
What distinguishes mammals is something else. Around one truly new acquisition, the mobile face, this old assemblage came into the service of a new function: not the saving of oxygen or the protection of the airway, but the showing, sounding, and reading of an inner state between conspecifics. The social engagement system is thereby not merely an example of exaptation. It is a whole ensemble of old parts and old connections, repurposed, expanded with a new element, and as a whole put into the service of something none of the parts could ever do separately: social connection.
How tightly this whole is neurally integrated, in the sense Porges has in mind, is a question that requires further research; about this the comparative physiology of Taylor and his colleagues makes no pronouncement. But that mammals possess a communication channel no other animal has and that this channel has carried the old heart regulation along into a new, social function is beyond doubt.
And it is precisely that communication channel that touches on what forms the heart 😉 of polyvagal theory: the concept of safety. For where this economical, delicate regulatory system really makes a difference is in the proximity of conspecifics. An animal that can choose only between sympathetic defense, fight or flight, or dorsal shutdown soon experiences the proximity of another as a threat. But an animal that can subtly fine-tune its heart rate and its facial expression via the ventral vagus can be near another without going on the defensive. It can tolerate proximity and even seek it out. That is physiologically important because it means that the proximity of a conspecific need not immediately turn into a fight-or-flight reaction. And there we also find the precondition for everything that subsequently became socially possible: not fighting but attuning, not fleeing but connecting, not merely surviving but surviving together. The ventral vagus did not bring about that cooperation on its own, because cooperation, which ultimately led to civilization, has many roots: from a prolonged childhood in which several adults care for one child, via shared language and intentionality, to the cultural capacity to pass on knowledge to following generations. But the ventral vagus did provide the bodily foundation on which a sense of safety in one another’s proximity could arise, and without that foundation the rest would not have been possible. An outdated mechanism, once at work in the regulation of the heart, thus became the silent precondition for a life among and with conspecifics.
What this means for theory and practice
This reformulation has consequences that are worth making explicit.
For polyvagal theory itself, the exaptation perspective means that the evolutionary claims as Porges originally formulated them can no longer serve as necessary underpinning. The statement that only mammals would have a myelinated vagus, or that the dorsal motor nucleus is evolutionarily older than the nucleus ambiguus, does not hold up against what current comparative physiology shows. Those statements therefore need no longer be part of the theoretical edifice. What remains, and what is well defensible empirically and clinically, is the claim that mammals, and specifically humans, have a coherent face-voice-heart system that couples social behavior and bodily regulation in a way that has no equivalent elsewhere in the animal kingdom. This claim does not stand or fall with the question of whether the nucleus ambiguus already occurs in lizards.
For clinical practice, this is a reassuring outcome. The usefulness of polyvagal-informed approaches (social contact as a regulator of physiological state, vagal tone as a marker of flexibility, co-regulation as a treatment element) is largely independent of the question of whether the critics or Porges are right about sharks and lungfishes. The observations that carry the clinical theory are observations of humans under social and more stressful circumstances. These remain valid as long as they are empirically confirmed in their own right.
And yet the distant past and the consulting room touch one another. What took shape over hundreds of millions of years, namely that proximity becomes possible only when the body feels safe enough to let down its defenses, is precisely what plays out on a small scale in every treatment room. Co-regulation works not in the first place through a technique, but because the other’s nervous system learns to register proximity as safe again. The therapist does, perhaps even in a single encounter, what the ventral vagus made possible over the course of evolution.
For the scientific discussion, finally, the exaptation perspective means that the war of words may come to rest. Taylor and his colleagues end their article with a remark that may be considered an invitation to Porges: a theory deserves regular testing, and they are curious how polyvagal theory will develop under that testing.
The question is not who has the right on their side. The question is whether polyvagal theory is willing to adjust its vocabulary in such a way that the building-block claim is detached from the coherence claim and thereby acquires an evolutionarily more precise and clinically stronger formulation.
How old?
So what is the answer to the question this article began with: how old is the ventral vagus, really?
That depends on what you mean by it. The anatomical building blocks, ventrolaterally located cardiac vagal neurons with myelinated fibers to the heart, are at least five hundred million years old and go back to the common ancestor of all vertebrates. The respiratory-modulated version of it, the pattern in which the heart rhythm fluctuates to the rhythm of breathing, is about four hundred million years old and is connected with the origin of lung breathing. The specific integration of heart, breathing, voice, and face into one coherent regulatory system around a mobile and expressive face is about two hundred million years old and coincides with the origin of mammals.
The ventral vagus is therefore at once ancient and very “young.” Which answer is correct depends on the level at which you look. And that is where the exaptation perspective shows its value.
In conclusion
The comparative physiology of Taylor, Wang and Leite is not an attack on polyvagal theory as a therapeutic framework. It is a correction of a specific evolutionary claim in a field where these three researchers are among the greatest authorities in the world. Anyone who reads their work as a rejection of PVT misses an opportunity for clarification. Anyone who reads it as building material for a more mature formulation of polyvagal theory, a formulation that distinguishes its building blocks from its coherence and explains that coherence as exaptation rather than as anatomical invention, holds in their hands a framework of thought that supports clinical practice better than the original version did.
A delimitation is in order here. The criticism discussed in this article is the evolutionary criticism: the question of whether the building blocks of the system are really specifically mammalian. That is not the only criticism polyvagal theory has received. There are also methodological questions, for example, about how ventral vagal activity can best be measured, and there is discussion about the clinical claims. These fall outside the scope of this article. What I have wanted to show here is that precisely the evolutionary objections, which at first glance seem to hit hardest, lose their sharpness as soon as you view them through the lens of exaptation. The building blocks are old; that point Taylor and colleagues score. What mammals do with them is new; that point remains for Porges. And exaptation is the key point at which those two truths come together.
A brief methodological remark to finish. Earlier in this article I noted that heart, voice, and face in mammals together do something the separate parts cannot, and I promised to come back to it. Anyone who thought of emergence at that point, I wrote, had been paying close attention. Emergence is the phenomenon whereby a coherent system displays properties that are not present in the separate parts. Anyone who reads polyvagal theory as a description of an integrated regulatory system sees in it an emergent layer: co-regulation, a sense of social safety, prosody, and affective attunement as properties that the whole system brings forth, not the separate parts.
One question finally remains hanging. In recent years Porges himself already shifted towards a language of repurposing. Given this movement, when would he himself have taken the step to the concept of exaptation?
Afterword
The exaptation perspective does, as I see it, three things at once in the polyvagal debate.
Firstly, it resolves a pseudo-opposition that has kept the discussion in a stalemate for years: the choice between mammalian uniqueness and evolutionary continuity. Both sides have been right, only at different levels, and the concept of exaptation provides the language to make that difference explicit.
Secondly, it shifts the heaviest claim of polyvagal theory from a vulnerable anatomical level, where Taylor and his colleagues stand empirically strong, to a synthesis level that is well defensible empirically. What is really organized differently in mammals is not the presence of separate building blocks, but the integration of face, voice, and heart into one coherent regulatory system. That claim does not stand or fall with the question whether certain vagal fibers already occur in the lungfish.
Thirdly, and this seems paradoxical but it is not, it actually makes polyvagal theory more testable than in its original form. The claims that remain are about what is measurable and researchable in humans: the degree to which face, voice, and heart respond to one another; the working of co-regulation; and the relationship between vagal tone and social behavior. These are empirical questions, not evolutionary claims that withdraw from testing.
What exaptation does not do, and I want to be honest about this, is remove all criticism of polyvagal theory. There are still open methodological questions, such as how ventral vagal activity can best be measured. And the specific phylogenetic claims around the evolutionary reorganization of vagal circuits are plausible but less firmly underpinned than the anatomical part of the theory. However, whether the picture of integration that Porges sketches is really realized neurally remains an open empirical question.
But for the evolutionary part of the debate, precisely where the PVT community has so far responded least convincingly, exaptation offers what the debate needs: a reformulation in which no winner or loser is designated, but both sides are reconciled at a higher level. And it is not without significance that Porges himself has, since 2021, been moving in that direction with his language of “repurposing.” He does not use the word exaptation, but he says something that lies very close to it. This article makes that movement explicit and places it within an evolutionary concept that biology has been working with since 1982.
One observation to finish. Perhaps the most lasting contribution of this whole exercise is not even the exaptation argument itself, but the three-layer structure that comes with it: evolutionary, neurobiological, and functional. That is no longer an exaptation-technical matter but a framework of thought that can also help, in other debates in the polyvagal literature, to disentangle confusions between levels. If anything from this piece is going to take on a life of its own in the discussion, I suspect it may lie there.
The old vagus is older than we thought,
but what mammals did with it was new.
If you found this article worth reading (you did, I hope?) and (not yet) feel like getting a paid subscription, you can always treat me to a cappuccino!
References
Berta, A., Sumich, J. L., & Kovacs, K. M. (2015). Marine mammals: Evolutionary biology (3rd ed.). Academic Press / Elsevier.
Campbell, H. A., Leite, C. A. C., Wang, T., Skals, M., Abe, A. S., Egginton, S., Rantin, F. T., Bishop, C. M., & Taylor, E. W. (2006). Evidence for a respiratory component, similar to mammalian respiratory sinus arrhythmia, in the heart rate variability signal from the rattlesnake, Crotalus durissus terrificus. Journal of Experimental Biology, 209(14), 2628–2636. https://doi.org/10.1242/jeb.02278
Doody, J. S., Burghardt, G. M., & Dinets, V. (2023). The evolution of sociality and the polyvagal theory. Biological Psychology, 180, 108569. https://doi.org/10.1016/j.biopsycho.2023.108569
Doody, J. S., Dinets, V., & Burghardt, G. M. (2021). The Secret Social Lives of Reptiles. Baltimore: Johns Hopkins University Press.
Gould, S. J., & Vrba, E. S. (1982). Exaptation, a missing term in the science of form. Paleobiology, 8(1), 4–15.
https://doi.org/10.1017/S0094837300004310
Grigg, G., Nowack, J., Bicudo, J. E. P. W., Bal, N. C., Woodward, H. N., & Seymour, R. S. (2021). Whole-body endothermy: Ancient, homologous and widespread among the ancestors of mammals, birds and crocodylians. Biological Reviews, 97(2), 766–801.
https://doi.org/10.1111/brv.12822
Grossman, P. (2023). Fundamental challenges and likely refutations of the five basic premises of the polyvagal theory. Biological Psychology, 180, 108589. https://doi.org/10.1016/j.biopsycho.2023.108589
Grossman, P., & Taylor, E. W. (2007). Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biological Psychology, 74(2), 263–285. https://doi.org/10.1016/j.biopsycho.2005.11.014
Halder, G., Callaerts, P., & Gehring, W. J. (1995). Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science, 267(5205), 1788–1792.
https://doi.org/10.1126/science.7892602
Popper, K. R. (1959). The Logic of Scientific Discovery. London: Hutchinson.
Porges, S. W. (1995). Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A Polyvagal Theory. Psychophysiology, 32(4), 301–318.
https://doi.org/10.1111/j.1469-8986.1995.tb01213.x
Porges, S. W. (2011). The Polyvagal Theory: Neurophysiological foundations of emotions, attachment, communication, and self-regulation. New York: W.W. Norton.
Porges, S. W. (2021). Polyvagal Theory: A biobehavioral journey to sociality. Comprehensive Psychoneuroendocrinology, 7, 100069. https://doi.org/10.1016/j.cpnec.2021.100069
Porges, S. W. (2022). Polyvagal Theory: A science of safety. Frontiers in Integrative Neuroscience, 16, 871227.
https://doi.org/10.3389/fnint.2022.871227
Porges, S. W. (2023). The vagal paradox: A polyvagal solution. Comprehensive Psychoneuroendocrinology, 16, 100200. https://doi.org/10.1016/j.cpnec.2023.100200
Shubin, N. (2008). Your Inner Fish. New York: Pantheon Books.
Taylor, E. W., Wang, T., & Leite, C. A. C. (2022). An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the ‘Polyvagal Theory’. Biological Psychology, 172, 108382. https://doi.org/10.1016/j.biopsycho.2022.108382
Just as with the word “branch” further on in the text, “pathway” here is a simplification. Strictly speaking, it is not a separate neural pathway but a bundle of fibers with its own origin and its own properties, running along within the larger bundle that is the vagus nerve. I use “pathway” here for the sake of readability, but keep in mind that it concerns fibers within one nerve, not a separate cable.
Taylor’s research in reality encompasses all classes of vertebrate animals (from sharks to mammals) and crustaceans besides. The enumeration in the main text confines itself to the groups that come up in this article. University profile: https://www.birmingham.ac.uk/staff/profiles/biosciences/taylor-edwin.
Besides DMNX we also regularly encounter the abbreviation DVN. They refer to the same brainstem nucleus, the dorsomedial motor nucleus of the vagus nerve. DMNX is the abbreviation as used in the polyvagal literature; DVN is the abbreviation accepted in current comparative physiology (Taylor et al., 2022). For alignment with Porges’s terminology I use DMNX in this article.
More concretely: the split between the lungfish line and the line that would eventually lead to mammals lies in the early Devonian, about 400 million years ago. The split between the amphibian line (from which toads emerged) and the amniote line (from which reptiles, birds and mammals arose) is younger, about 340 million years ago. Both splits therefore lie deep in the Paleozoic, well before the appearance of the first true mammals, which was the case around 200 million years ago.
Strictly speaking it is somewhat more complicated. Amphibians have a heart with two atria and one ventricle, in which oxygen-poor and oxygen-rich blood partly mix in the same chamber. Most reptiles (turtles, lizards, snakes) also have one ventricle, internally partly divided by an incomplete septum. What many people do not know: almost all reptiles have not one but two aortas, a left and a right, alongside the pulmonary artery. Unlike us, that is.
Crocodilians form a fascinating exception within the reptiles. They do have two fully separated ventricles, just like birds and mammals, but they retained the reptile-specific two-aorta system. In them the right aorta arises from the left ventricle (as is to be expected; this aorta follows a course comparable to that in humans), but the left aorta arises from the right ventricle (which is unusual, but that goes too deep for this article). At the point where these two aortas pass each other at the base of the heart there is an opening called the foramen of Panizza. Through it, blood can under certain circumstances still cross between the two aortas, and oxygen-poor blood can be sent to the body via the left aorta without first passing through the lungs. That is a beautiful evolutionary functional feature: while diving, crocodiles can effectively bypass their pulmonary circulation to save oxygen.
For the mechanism discussed in this article, the decisive point is not the number of ventricles, but whether blood can under certain circumstances be diverted between the pulmonary and systemic circulations. In fishes, amphibians and almost all reptiles this is possible. In mammals and birds it is not. Crocodiles in this respect show something beautiful: they demonstrate that anatomical separations and functional separations are not the same thing. Two ventricles does not automatically mean that the circulation is fully separated.
When researchers expressed the eyeless gene (and its mammalian counterpart Pax6) at an aberrant location in a fruit fly, such as on a leg, wing or antenna, a complete fly eye grew there (Halder, Callaerts & Gehring, 1995). The switch gene therefore gave the starting signal “build an eye here”, and the fly carried that out with its own genetic tools. The effect is so deeply conserved that even the Pax6 gene of a mouse or a squid achieves this in a fly, despite these animal groups having diverged hundreds of millions of years ago. The structure of the eye is therefore invented again and again, but the genetic toolkit with which that happens is ancient and shared. With this, the eye illustrates on a small scale precisely what this article argues on a larger scale: old building blocks that are deployed again and again for comparable purposes.
The concept of exaptation has been well established in evolutionary thinking since the article by Stephen Jay Gould and Elisabeth Vrba in 1982. A possible objection is that I am deploying exaptation post hoc in my article in order to rescue a theory. That would misjudge the concept: exaptation is standard tooling in evolutionary biology for understanding functional shifts, not an ad hoc repair.
It really is running the gauntlet when it comes to formulating! I had first written: “The core of exaptation is that the current function of a trait need not be the reason why that trait once arose.” A reason, however, soon implies a purpose, and the whole of evolution has no purpose, no preconceived plan. In a few places in the article I will surely be unable to avoid an impure formulation, but then again, the readability of an article also asks something.
This second criterion, recognizing a trait by the traces of its past, is beautifully illustrated by Stephen Jay Gould in his famous essay on the panda’s thumb. The giant panda has a “thumb” with which it holds bamboo, but that thumb is not a real finger. It is an enlarged wrist bone that has acquired a new function. Precisely the fact that the construction comes across as clumsy and improvised betrays that it was not designed for gripping, but is a repurposed little bone that happened to come in handy. Gould used this to show that imperfection is often better evidence for evolution than perfection: a perfectly fitting trait could, after all, also have been designed, but a clumsy makeshift solution betrays a history of reuse. With the vagus we see something comparable. That the “breath-rhythmic” fluctuation of the heartbeat in mammals no longer has a clear physiological function is precisely such a trace: it looks like a remnant, not like a purpose-built, fine-tuned adaptation.
Empirical means: based on observation and experience, on what you can establish in reality, instead of on reasoning, assumption or conviction alone. An empirical question is a question you answer by looking, measuring and comparing, not by thinking about what would be logical or desirable. The word comes from the Greek empeiria, which means experience.
Anyone who now thinks of the concept of “emergence” has been paying close attention; I come back to it at the end.
Whales and dolphins form an interesting exception here within the mammals. When they returned to the water, they largely lost their facial musculature, but the nucleus ambiguus, the central nucleus of the ventral vagal complex, they retained as mammalian heritage. Their social co-regulation runs mainly via vocalizations and bodily contact. That is anatomically consistent with the idea that the phylogenetic architecture of the VVC is preserved, even when the specific elaboration differs from one mammal group to another (Berta, Sumich & Kovacs, 2015).
Porges’s formulation is unfortunately chosen and thereby vulnerable to attack: critics read into it a wish not to be falsifiable, especially because often only the first part of his statement is quoted, namely “The theory was not intended to be proven or refuted”! If you read the statement in its entirety, namely “The theory was not intended to be proven or refuted, but to be informed by research and modified”, then you read a more defensible intention, namely that his theory is rather a broad organizing framework than a single testable hypothesis. That distinction is common in the philosophy of science: an overarching framework is rarely confirmed or refuted in its entirety, but it does yield testable sub-hypotheses. The unfortunate thing therefore lies not in what Porges presumably means, but in how he formulates it.
A testable design could be the following: have two people enter into conversation with each other and simultaneously measure their facial expressions, the timbre and melody of their voice, and their heart rate variability (HRV). If face, voice and heart in mammals really form one regulatory system, then changes in these three domains would not vary independently of one another, and a warmer voice and friendlier facial expression in the one would evoke a measurable change in the HRV of the other, which we call co-regulation. One way to falsify the claim runs via people with Möbius syndrome, a congenital condition in which the facial muscles do not work: if in them the co-regulation via voice and heartbeat remains fully intact, then the claim that the face is a necessary link comes under pressure. Other studies can likewise be conceived.