

Hoe oud is die ventrale vagus eigenlijk?
Ouder dan Porges vermoedde, slimmer dan Taylor benoemt. Het ontwarren van een wetenschappelijk debat over de polyvagaaltheorie


Do you prefer the English version?
Welkom bij mijn langste artikel op RelaxMore.net tot nu toe. Het heeft me behoorlijk wat werk gekost, maar het was de moeite waard. Het doornemen van de wetenschappelijke bronnen was intensief, leerzaam en leuk. Maar het meeste werk zat in het zoeken naar de formulering die recht doet aan de beide kanten van een debat dat al bijna dertig jaar duurt en waar ik nu over schrijf. Neem gerust de tijd om het te lezen. Omdat het een lang stuk is, begin ik met een samenvatting. Ik hoop ook dat je daarna des te nieuwsgieriger bent geworden naar hoe het verhaal zich ontvouwt.
Samenvatting
Sinds halverwege de jaren negentig wordt er gediscussieerd over het evolutionaire fundament van de polyvagaaltheorie. Aan de ene kant Stephen Porges, die het ventraal vagale systeem (= het sociale verbindingssysteem) presenteert als typisch zoogdier-eigen: een nieuwe, snelle vagale rem op het hart, vanuit de nucleus ambiguus, die zoogdieren in staat stelt om sociaal contact te zoeken en elkaar te kalmeren. Aan de andere kant een groep vergelijkende fysiologen onder leiding van Edwin Taylor, die heeft aangetoond dat de bouwstenen die Porges noemt – gemyeliniseerde cardiale vagale vezels, twee kernen in de hersenstam en een respiratoir gemoduleerd hartritme (ik leg alle termen verderop uit) – al honderden miljoenen jaren vóór de zoogdieren bestonden, bij haaien, longvissen, kikkers en reptielen.
Op het eerste oog lijkt dit een discussie waar een winnaar uit moet komen. Maar de tegenstelling blijkt niet meer zo spectaculair zodra je het evolutionaire concept exaptatie gaat toepassen: het verschijnsel dat een bestaande structuur in een nieuwe context een nieuwe functie krijgt. De bouwstenen zijn dus oud, maar wat zoogdieren ermee hebben gedaan, is wel nieuw: een integratie van hart, ademhaling en stem in één samenhangend regulatiesysteem, georganiseerd rondom iets wat geen ander gewerveld dier in deze mate bezit, een beweeglijk en expressief gezicht.
In dit artikel laat ik zien hoe dat perspectief Porges en zijn critici binnen één evolutionair kader verenigt, en hoe het de polyvagaaltheorie, paradoxaal genoeg, juist empirisch toetsbaarder maakt dan in haar oorspronkelijke vorm.
Een meningsverschil van dertig jaar
Sinds halverwege de jaren negentig wordt er gediscussieerd over het evolutionaire fundament van de polyvagaaltheorie (Porges, 1995, 2011). Aan de ene kant staat Stephen Porges, de Amerikaanse wetenschapper die de theorie heeft ontwikkeld. Hij stelt dat zoogdieren, en dus ook wij mensen, beschikken over een speciale zenuwbaan die andere dieren niet hebben. Deze zenuwbaan1 is onderdeel van de tiende hersenzenuw, de bekende nervus vagus. Volgens Porges maakt deze baan het mogelijk dat zoogdieren rust kunnen vinden bij elkaar en veiligheid aan elkaar kunnen geven met hun lichaamstaal, stem en mimiek. Het is het neurale, lichamelijke en gedragsmatige fundament van wat hij het social engagementsystem, het sociale verbindingssysteem noemt.
Aan de andere kant staat een groep vergelijkende fysiologen, onderzoekers die zenuwstelsels van verschillende diergroepen met elkaar vergelijken. De Britse emeritus hoogleraar Edwin Taylor (University of Birmingham) doet samen met Tobias Wang (Aarhus University) en Cleo Leite (Federal University of São Carlos) al decennia onderzoek naar hartslag- en ademhalingsregulatie en de coördinatie daarvan bij vissen, kikkers, hagedissen en vogels2. Zij publiceerden in 2022 een artikel met de titel “An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the ‘Polyvagal Theory’”, in het Nederlands “Een overzicht van de fylogenie (= de studie van de afstammingsgeschiedenis van een groep organismen) van cardiorespiratoire regulatie (= de regulatie van hartslag en ademhaling) bij gewervelden, met enkele beschouwingen over de ‘Polyvagaaltheorie’”.
Hun bezwaar tegen de polyvagaaltheorie is niet dat het functionele verhaal van Porges over sociaal gedrag onzinnig zou zijn. Hun bezwaar zit dieper: zij zeggen dat de bouwstenen waar Porges het systeem op baseert – in tegenstelling tot wat hij beweert – helemaal niet exclusief bij zoogdieren voorkomen. Die zenuwvezels, die hersenstamkernen, dat samenspel tussen ademen en hartslag: dat vind je bij vissen, hagedissen en kikkers ook al. Taylor, Wang en Leite beschrijven dit standpunt uitgebreid in hun, overigens zeer interessante, artikel (Taylor, Wang, & Leite, 2022, zie hierboven voor een pdf). Eerder was Paul Grossman al met een methodologische kritiek gekomen die in vergelijkbare richting wees (Grossman & Taylor, 2007; Grossman, 2023).
Op het eerste oog lijkt dit een gevecht waar maar één winnaar uit kan komen. Of de bouwstenen zijn typisch voor zoogdieren, of dat zijn ze niet. Maar wie het debat op de of-of-manier voert, mist iets belangrijks. Er is een derde mogelijkheid die beide kanten recht doet, een en-en-model dus. De mogelijkheid daarvoor ligt verborgen in een evolutionair concept dat exaptatie heet, een woord dat ik verderop uitleg. Met behulp van dat concept blijkt de tegenstelling minder onverzoenlijk dan ze lijkt.

We beginnen bij de basis
Voordat we het debat zelf induiken, zal ik eerst een paar begrippen uitleggen, zodat je de redenering kunt volgen.
Nervus vagus
De nervus vagus is de tiende hersenzenuw. Het is een lange, best dikke zenuw die vanuit de hersenstam naar beneden loopt en zich vertakt naar het hart, de longen, het strottenhoofd en de organen in de buik. Het is de langste zenuw in het lichaam. De vagus hoort bij het parasympathische zenuwstelsel: het deel van ons zenuwstelsel dat actief is in rust en herstel, dat de hartslag vertraagt, de spijsvertering ondersteunt, en betrokken is bij stem en gezichtsuitdrukking, maar ook bij shutdown- en verslappingsresponsen bij stress die we als levensbedreigend ervaren.
De vagus heeft niet één oorsprong in de hersenstam, maar twee. In de hersenstam is een groepje zenuwcellen meer naar de rugzijde gelegen: de dorsale (= achterzijde) motorkern van de vagus, afgekort de DMNX. En er is een groepje cellen meer naar de buikzijde (= ventraal) gelegen: de nucleus ambiguus, afgekort de NAmb. Dat het om twee verschillende hersenstamkernen gaat, is belangrijk in de polyvagaaltheorie. Porges koppelt ze namelijk aan verschillende functies. De DMNX is volgens hem evolutionair oud en hangt samen met ineenstorten, dichtklappen en dissociatie bij overweldigende dreiging, wat in de polyvagaaltheorie shutdown of collapse heet. De NAmb daarentegen zou bij zoogdieren een functie hebben die geen andere diergroep kent. Niet zozeer omdat deze kern elders zou ontbreken (Porges erkent zelf dat een NAmb-achtige structuur al bij reptielen aanwezig is), maar omdat de snelle, gemyeliniseerde vezels (dat is het volgende begrip dat ik uitleg) die vanuit de NAmb naar het hart lopen, en de manier waarop ze met gezicht, stem en gehoor zijn verbonden, volgens hem uniek voor zoogdieren zouden zijn. Die NAmb-tak zou het mogelijk maken om sociaal contact te zoeken, te kalmeren en in verbinding te zijn.
Een waarschuwing is hier op zijn plaats, want ik gebruikte zojuist het woord “tak”, en dat woord is eigenlijk misleidend. Je komt het overal tegen in de literatuur over de polyvagaaltheorie: de ventrale vagale tak, de dorsale vagale tak, alsof het om twee aparte kabels gaat die ieder hun eigen weg door het lichaam zoeken. Zo werkt het niet. De vezels die uit de DMNX komen en de vezels die uit de NAmb komen, lopen niet gescheiden, maar raken al voor het verlaten van de hersenstam vervlochten in dezelfde zenuwstreng. Stel je dus geen twee losse draden voor, maar één touw waarin draden van verschillende herkomst samen zijn gevlochten. De cardiale tak van de vagus, die naar het hart loopt, bevat dan ook vezels uit beide kernen, snelle gemyeliniseerde én trage ongemyeliniseerde, door elkaar heen (Taylor et al., 2022). Ik blijf in dit artikel overigens gewoon over "takken" spreken, want het is een handig en ingeburgerd woord, maar je weet nu dat het een vereenvoudiging is.
Dat de twee soorten vezels functioneel te onderscheiden zijn, betekent dus niet dat ze anatomisch gescheiden zijn. Het onderscheid zit in hun oorsprong en in hun gedrag, niet in twee losse takken. Dit lijkt een detail, maar het raakt aan iets dat in dit hele artikel telkens terugkomt: een functioneel onderscheid is iets anders dan een fysieke scheiding. Wie de ventrale en de dorsale vagus als twee aparte zenuwbanen voorstelt, maakt precies de denkfout die ook ten grondslag ligt aan een deel van de kritiek op de polyvagaaltheorie. Ik kom daar verderop op terug.
Myeline
Nog één begrip is van belang: myeline. Sommige zenuwvezels zijn omhuld door een isolatielaagje van vet en eiwit. Dat laagje, myeline, zorgt ervoor dat signalen veel sneller door de vezels kunnen reizen. Vergelijk het met de plastic isolatie om een elektriciteitsdraad. Zenuwvezels mét myeline geleiden tot wel 10 keer sneller dan vezels zonder myeline. Een belangrijke stelling van Porges over de aansturing van het hart is dat alleen zoogdieren daarvoor beschikken over gemyeliniseerde NAmb-vezels, en dat deze snelle vezels de basis vormen voor het fijngevoelige samenspel tussen ademen en hartslag dat zich uit in de hartslagvariabiliteit.
Dat samenspel zelf heet respiratoire sinusaritmie, afgekort RSA. Het is het natuurlijke verschijnsel dat het hart bij inademen iets sneller gaat en bij uitademen iets vertraagt. Het wordt vaak gezien als een teken van een gezond, flexibel zenuwstelsel.
Wat Taylor en collega’s hebben aangetoond
Met deze begrippen op zak kunnen we kijken naar wat Taylor, Wang en Leite in 2022 in het tijdschrift Biological Psychology hebben gepubliceerd (Taylor et al., 2022). Hun overzichtsartikel vat decennia onderzoek samen tot een gestructureerde kritiek op de evolutionaire onderbouwing van de polyvagaaltheorie. Hun argumentatie heeft vier hoofdlijnen.
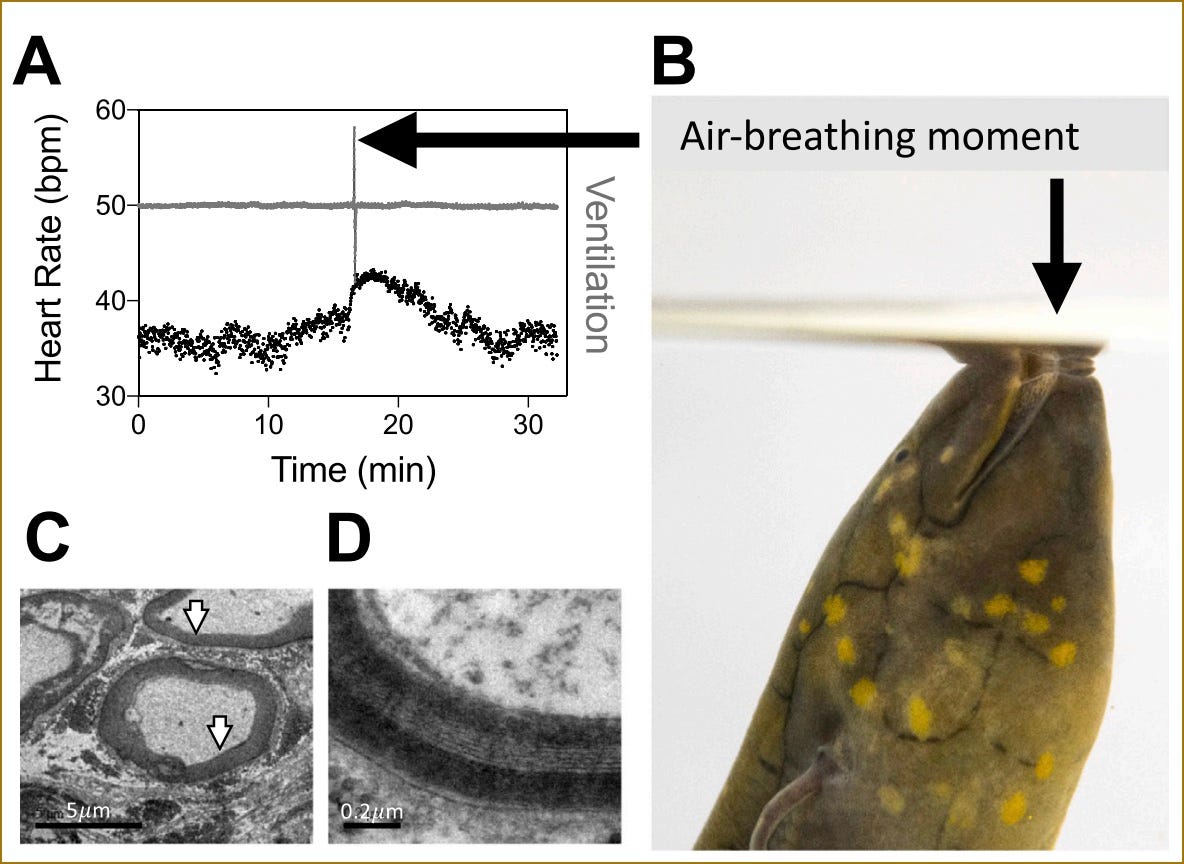
De eerste lijn betreft de remming van het hart in rust. Bij alle bestudeerde gewervelde dieren, van haaien tot zoogdieren, blijkt dat de hartslag in rust wordt afgeremd door de vagus. Bij reptielen blijkt deze rem zelfs heel sterk te zijn. Bij de ratelslang verdubbelt de hartslag ongeveer wanneer de vagale invloed met atropine wordt weggenomen (Campbell et al., 2006), terwijl de stijging bij de mens doorgaans tussen de dertig en vijftig procent blijft. Een traag metabolisme gaat dus prima samen met een krachtige vagale grip op het hart. Dat is even wennen voor de intuïtie, die van een koudbloedig, traag dier eerder een losse dan een strakke teugel verwacht. Sterke vagale controle van het hart in rust is hoe dan ook dus geen zoogdiernieuwtje. Of die controle bij reptielen ook berust op dezelfde snelle, gemyeliniseerde vezels als bij ons, is een aparte vraag, waarop ik bij de derde lijn terugkom.
De tweede lijn betreft de twee oorsprongen van de vagus in de hersenstam. Porges presenteert het als een typisch zoogdier-kenmerk dat er vagusvezels die het hart aansturen ontspringen uit twee kernen in de hersenstam: in de DMNX én in de NAmb. Maar Taylor en zijn collega’s laten zien dat deze tweedeling al bestaat bij de hondshaai. Bij deze haai zit het grootste deel van de vagale zenuwcellen in de DMNX, maar er is óók een groepje meer naar voren (= ventraal) gelegen, en in dat groepje liggen al cellen die het hart aansturen. Taylor en collega's zien deze ventrolaterale groep als een mogelijke voorloper van wat bij zoogdieren de nucleus ambiguus is geworden. Bij krokodillen, hagedissen, slangen en kikkers vinden de auteurs vergelijkbare verdelingen. Bij de Afrikaanse klauwkikker ligt zelfs 30% van de betreffende vagale zenuwcellen al in die voorste hersenstamkern. De tweedeling is niet alleen oud, in het ventrale groepje liggen bij de haai al cardiale vagale neuronen, op een locatie die door Taylor als een mogelijke voorloper van de zoogdier-NAmb wordt gezien. En de relatieve omvang ervan loopt door de evolutie heen op, van tien procent bij de haai naar dertig bij de klauwkikker en een uitgebreide kern bij ons. Geen uitvinding van ‘ons’ dus, maar een uitbouw.
De derde lijn betreft de gemyeliniseerde vezels. De claim van Porges is hier specifiek: niet dat er ergens in de vagus myeline aanwezig zou zijn, maar dat alleen zoogdieren snelle, gemyeliniseerde cardiale vezels bezitten die in een respiratoir-gemoduleerd (= door de ademhaling gestuurd) circuit het hart van moment tot moment kunnen bijsturen. Letterlijk: “Alleen zoogdieren hebben een gemyeliniseerde vagus” (Porges, 2009, 2011). Met de elektronenmicroscoop hebben Taylor en collega's deze vezels intussen ook in de cardiale vagustak aangetoond bij haaien, longvissen, ratelslangen, hagedissen en padden. Bij de hondshaai en de Zuid-Amerikaanse longvis blijken de geleidingssnelheden bovendien te matchen met die van bepaalde vezels bij zoogdieren. En zowel bij de longvis als bij de ratelslang draaien deze vezels mee in een respiratoir-gemoduleerd hartritme dat lijkt op de RSA die we bij zoogdieren kennen. Het gaat dus niet om enkele uitzonderingen waar ‘ergens’ myeline is aangetroffen, maar om hetzelfde functionele pakket dat Porges als zoogdier-eigen presenteert.
De vierde en theoretisch interessantste lijn betreft het samenspel tussen ademen en hartslag zelf. Taylor en zijn collega’s noemen dit cardiorespiratoire synchronisatie (CRS): elke hartslag valt samen met een ademhalingsbeweging, een 1-op-1 koppeling, aangestuurd vanuit de DMNX3, die de tegenstroom van bloed en water over de kieuwen optimaliseert voor maximale zuurstofopname. Bij luchtademhalers, zoals longvissen, kikkers, reptielen, vogels en zoogdieren, ziet de koppeling tussen ademhaling en hartslag er fundamenteel anders uit. De hartslag is dan veel sneller dan het ademritme, dus een strikte 1-op-1 koppeling is niet meer mogelijk. In plaats daarvan beïnvloedt de ademhaling de snelheid van de hartslag: tijdens inademing wordt de vagale rem op het hart even losgelaten, waardoor de hartslag versnelt, en bij uitademing gaat de rem er weer meer op. Dit patroon, waarin de hartslagsnelheid schommelt op het ritme van de ademhaling, is wat we bij zoogdieren respiratoire sinusaritmie (RSA) noemen. Ditzelfde mechanisme is dus aangetroffen bij longvissen en cururu-padden, dieren waarvan de afstammingslijn zich honderden miljoenen jaren geleden heeft afgesplitst van de lijn die uiteindelijk tot zoogdieren zou leiden4.

Het meest verrassende inzicht zit in de slotgedachte van de auteurs. Bij reptielen en amfibieën, die een hart hebben waarbij long- en lichaamscirculatie niet volledig gescheiden zijn – zoals bij ons het geval is – helpt deze schommeling in hartslag bij elke ademteug om bloed efficiënt naar de longen te sturen5. Zoogdieren hebben die efficiëntiewinst niet meer nodig: ons hart heeft volledig gescheiden boezems en kamers: voor de longcirculatie en voor de lichaamscirculatie. RSA bij zoogdieren is dus eigenlijk geen evolutionaire innovatie. Het is een overblijfsel uit een tijd waarin het ritme nog werkelijk werk te doen had, en nu nog altijd klopt😉, maar zonder de duidelijke fysiologische rol die het bij dieren met cardiale shunting nog vervult.
Hier kan een vraag opkomen. Als RSA bij zoogdieren zijn oorspronkelijke functie heeft verloren, waarom geldt hartslagvariabiliteit (waarvan RSA een belangrijk onderdeel is) dan als een maat voor gezondheid? Het antwoord is dat RSA bij ons niet waardevol is om wat het doet, maar om wat het verraadt. De schommeling van de hartslag met het ritme van de adem ontstaat doordat de snelle ventraal vagale vezels het hart voortdurend bijsturen. De omvang van die schommeling is daarmee een venster op hoe soepel die vagale regulatie werkt. Veel variabiliteit wijst op een flexibel, responsief systeem; weinig variabiliteit op een systeem dat hapert door stress, ziekte of ouderdom. RSA is bij zoogdieren dus geen functioneel mechanisme meer, maar een bruikbaar en meetbaar signaal, en in die hoedanigheid waardevol.
Wat Taylor nadrukkelijk níet beweert
In het artikel van Taylor en collega’s staan enkele opmerkingen die vaak onderbelicht blijven. De auteurs zeggen in hun inleiding en in hun conclusie expliciet dat ze niets willen zeggen over de rol van de vagus bij sociaal gedrag van mensen. Letterlijk schrijven ze dat ze niet pretenderen “iets nuttigs te kunnen zeggen over de mogelijke rollen van een social engagementsystem bij de mens, dat op een of andere wijze parasympathische input via de nervus vagus zou kunnen omvatten” (Taylor et al., 2022). Tegelijk laten ze er geen twijfel over bestaan dat ze het ‘smart vagus’-concept zelf, waarmee Porges dat sociaal engagementsysteem fundeert, als overbodig beschouwen. Hun afbakening betreft dus alleen de psychologische en klinische uitwerking van de theorie; over de neurobiologische bouwstenen daaronder spreken ze zich juist wél uit. Ze erkennen dat de vagus vele belangrijke functies vervult bij ons. Hun kritiek beperkt zich tot wat zij zelf bestudeerd hebben: de vergelijkende fysiologie van het hart en de ademhaling bij verschillende diergroepen.
Dit is een wezenlijke begrenzing van hun onderzoeksgebied, en wetenschappelijk zeer integer. Wat Taylor en zijn collega’s weerleggen, is de bewering dat de anatomische bouwstenen typisch zijn voor zoogdieren. Wat zij dus niet weerspreken, is de bewering dat zoogdieren een geïntegreerd systeem hebben waarin gezicht, stem en hart samenwerken om sociaal contact mogelijk te maken. Hier blijkt al dat de polyvagaaltheorie en de kritiek van Taylor en zijn collega’s elkaar niet hoeven uit te sluiten.
Longvis?
Op dit punt zul je je als nuchtere lezer misschien afvragen: wat heeft een longvis met mij te maken? Mijn ventrale vagus reguleert mijn omgang met collega’s, mijn vermogen om mijn kind te kalmeren, mijn stem in een gesprek. Een longvis flirt niet, glimlacht niet, troost geen partner. Wat doet die hier dan in het verhaal?
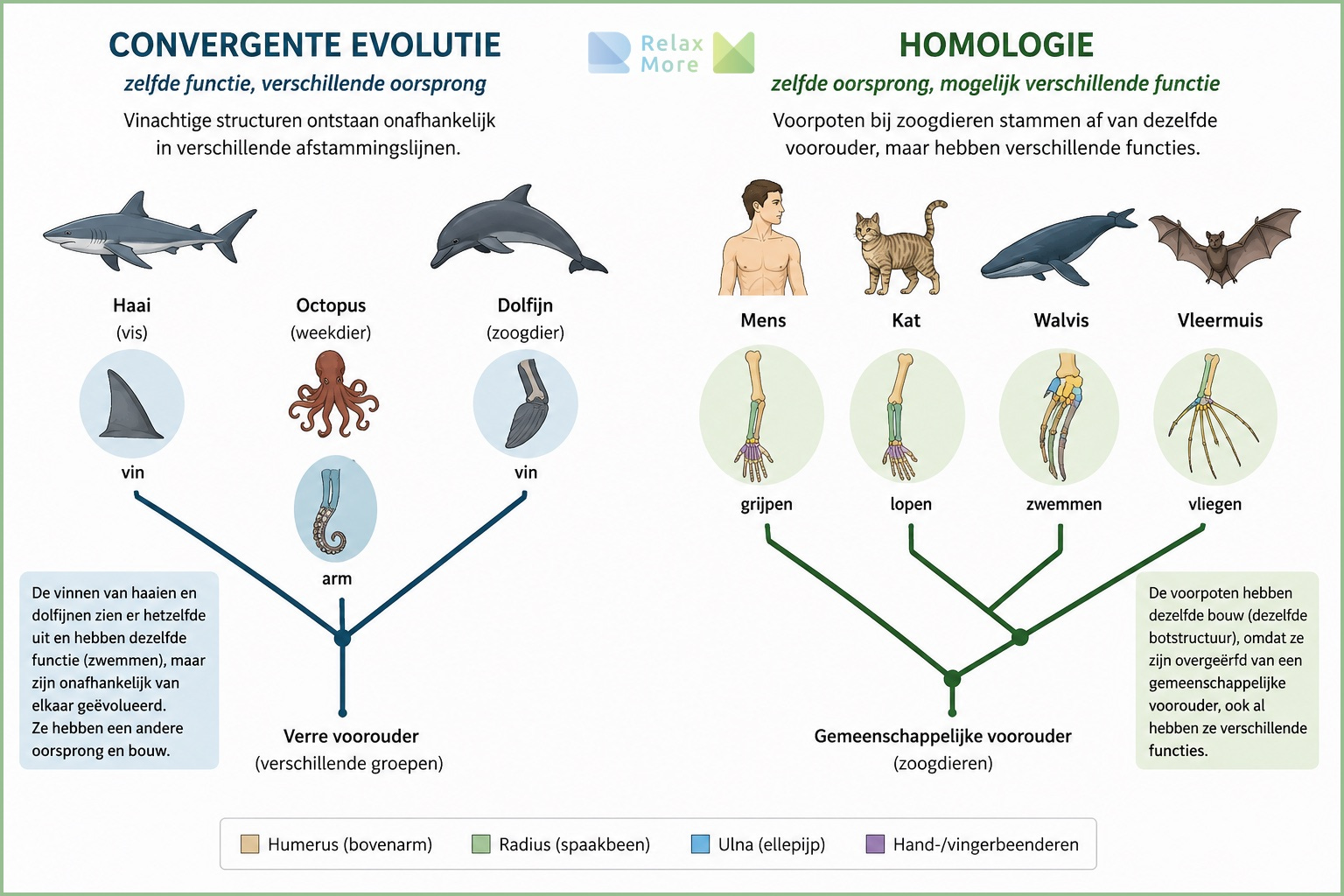
Het antwoord ligt in een onderscheid dat in de evolutionaire biologie van belang is. Wanneer een kenmerk in verschillende afstammingslijnen voorkomt, kan dat op twee fundamenteel verschillende manieren zijn ontstaan. Het kan onafhankelijk zijn uitgevonden, telkens opnieuw, in elke lijn apart. Dat heet convergente evolutie: vleermuizen, vogels en pterosauriërs hebben alle drie vleugels, maar ieder vanuit een eigen evolutionair pad. De gemeenschappelijke voorouder had geen vleugels. Of het kan geërfd zijn van een gemeenschappelijke voorouder die het kenmerk al bezat, waarna alle afstammingslijnen het hebben behouden. Dat heet homologie.
Zoals je hierboven kunt zien, lijkt het verschil tussen convergent en homoloog goed af te bakenen, maar in de praktijk lopen de twee nog wel eens door elkaar. Neem het oog. Naar schatting is een lichtgevoelig orgaan tientallen keren onafhankelijk van elkaar ontstaan in de dierenwereld, van het samengestelde oog van insecten tot het cameraoog van gewervelden en inktvissen. Op het niveau van de bouw is het oog dus een schoolvoorbeeld van convergentie: telkens opnieuw uitgevonden, met verschillende onderdelen en verschillende optica. En toch zit daar een verrassing onder. Al die onafhankelijk ontstane ogen blijken te worden aangezet door hetzelfde genetische schakelmechanisme: het zogeheten Pax6-gen speelt bij vrijwel alle dieren een sleutelrol in de aanleg van ogen. Dat gen is wél homoloog, geërfd van een verre gemeenschappelijke voorouder6.
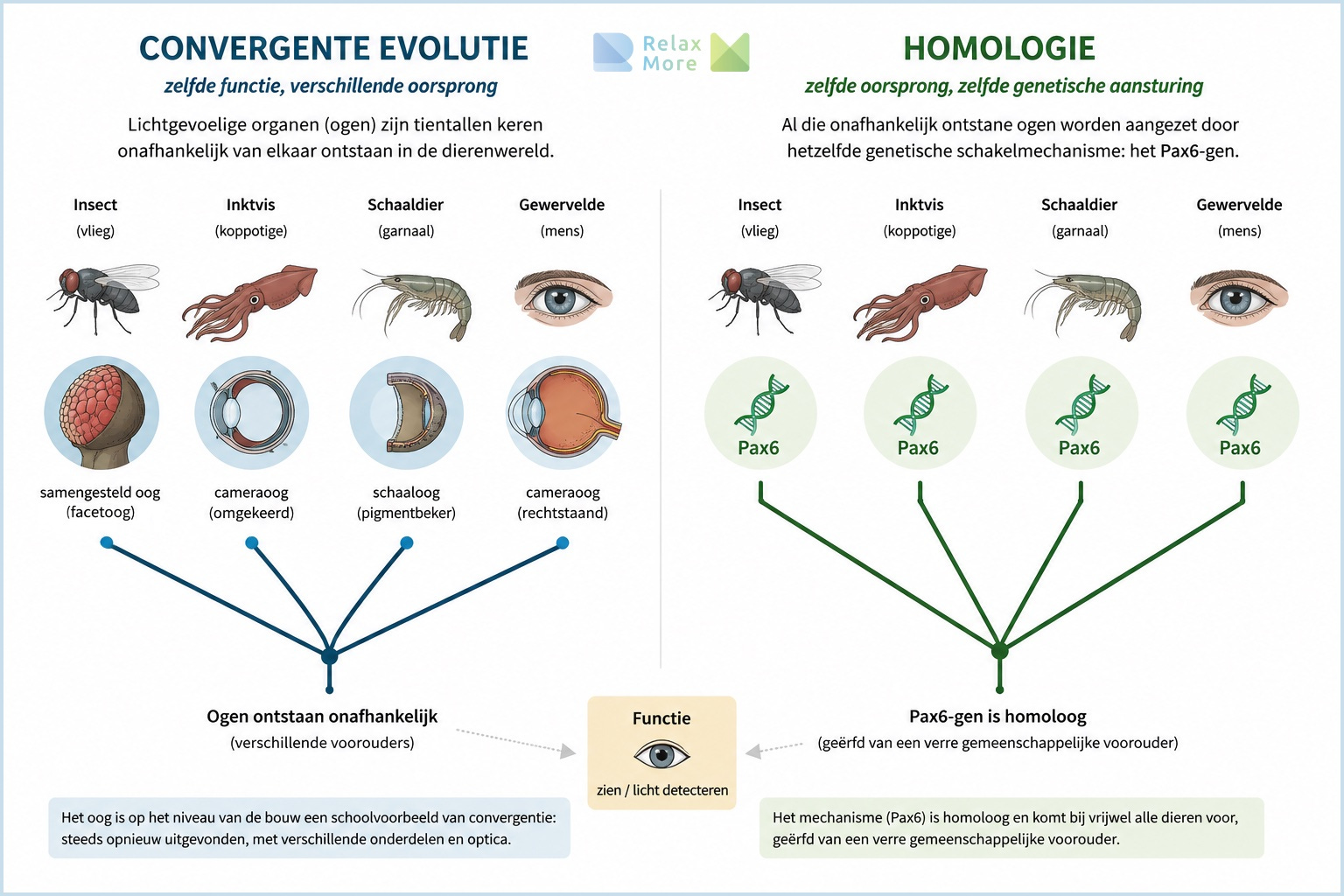
Het oog is dus tegelijk convergent en homoloog: convergent in zijn bouw, homoloog in zijn genetische aansturing. Het oude gen gaf bij verschillende dieren het startsein “bouw hier een oog”, en elke afstammingslijn vulde dat commando in met zijn eigen middelen. Precies dit verschil tussen niveaus is wat we nodig hebben om het vagale verhaal te begrijpen. Want ook daar geldt: je moet weten op welk niveau je kijkt voordat je kunt zeggen of iets oud of nieuw is.
Taylor en zijn collega’s nemen een heel duidelijke positie in: het mechanisme van de vagale rem op het hart, met gemyeliniseerde vezels en respiratoire modulatie, is homoloog. Het is niet onafhankelijk uitgevonden door longvissen, padden en zoogdieren, maar geërfd van een gemeenschappelijke voorouder die ergens in het Devoon leefde, ongeveer vierhonderd miljoen jaar geleden. Die voorouder had het mechanisme al: de snelle vezels, de cellen die het hart aansturen, en de koppeling met het ademritme. En al zijn afstammelingen, ieder via zijn eigen evolutionaire traject, hebben het meegekregen en in licht aangepaste vorm doorgegeven.
Dit is een belangrijker punt dan je misschien op het eerste gezicht zou zeggen. Als het kenmerk convergent was ontstaan, zou Porges nog kunnen volhouden dat zoogdieren het opnieuw hebben ontwikkeld, als unieke uitvinding op hun eigen evolutionaire pad. Maar homologie betekent dat het oud erfgoed is. De anatomische bouwstenen, een ventrale groep cardiale vagale neuronen met gemyeliniseerde vezels naar het hart, zijn niet typisch zoogdier-eigen. Ze zijn in herkenbare vorm al aanwezig bij de hondshaai en keren terug bij vrijwel alle latere gewervelden. En de RSA-achtige koppeling van hartritme en ademhaling die zoogdieren laten zien, delen ze met longvissen, kikkers, reptielen en vogels. Het mechanisme is dus niet nieuw bij zoogdieren; het is heel oud.
En hier raakt het dus wel degelijk de mens. Niet omdat wij iets met de longvis te maken hebben in ons dagelijkse leven, maar omdat het de tijdlijn van Porges’ theorie ondergraaft. Porges baseert zijn theorie op de claim dat zoogdieren een nieuwe bouwsteen hebben ontwikkeld die het sociale betrokkenheidssysteem mogelijk maakt. Als die bouwsteen er bij de longvis ook al was, is dat verhaal niet meer houdbaar in zijn oorspronkelijke vorm.
Het wordt dan tijd om opnieuw te bedenken wat zoogdieren werkelijk nieuw doen.
En dat is precies waar ik met dit hele artikel naartoe werk: niet de bouwstenen, maar wat zoogdieren ermee hebben gedaan, is nieuw. De longvis hoort dus thuis in dit verhaal, als getuige van een oude erfenis die wij met hem delen. De vraag wordt dan: hoe kunnen we begrijpen wat zoogdieren met die erfenis hebben gedaan?
Daar komt een evolutionair concept om de hoek kijken dat exaptatie heet.

Exaptatie
In 1982 publiceerden de Amerikaanse evolutionair bioloog Stephen Jay Gould en zijn collega Elisabeth Vrba een kort maar invloedrijk artikel onder de titel: ‘Exaptatie, een ontbrekende term in de wetenschap van de vorm’ (Gould & Vrba, 1982). Hun stelling was dat de evolutiebiologie tot dan toe een belangrijke categorie van vorm en functie miste.


Tot dan toe sprak men namelijk vooral over adaptaties: eigenschappen die door natuurlijke selectie zijn ontstaan ten behoeve van de functie die ze vervullen. De lange poten van een paard zijn een adaptatie om snel te kunnen rennen; de zwemvliezen van een eend zijn een adaptatie om te kunnen zwemmen. Maar Gould en Vrba wezen erop dat veel eigenschappen die nu een bepaalde functie vervullen, niet ten behoeve van die functie zijn ontstaan. Ze zijn in een andere omgeving, voor een ander doel gevormd, en gingen pas later een nieuwe functie vervullen. Voor dit type vorm-en-functie bedachten zij de term exaptatie7.
De term exaptatie is gevormd naar analogie van adaptatie. Waar het voorvoegsel ad- (= naartoe) wijst op een eigenschap die naar een doel toe is gevormd, wijst ex- (= vanuit) op een eigenschap die ergens anders vandaan komt en pas later van pas bleek te komen.
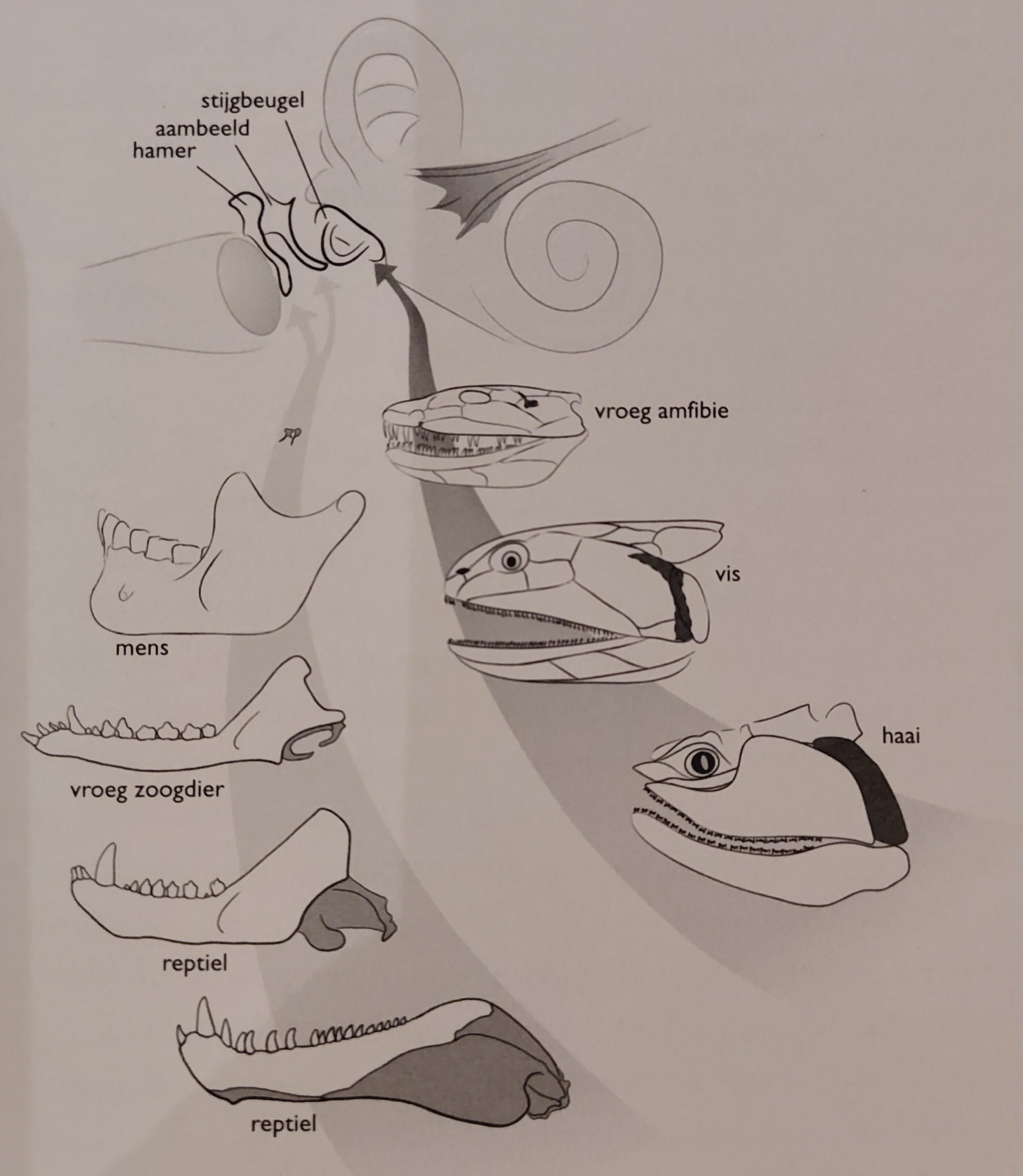
Enkele voorbeelden zijn waarschijnlijk verhelderend: vogelveren zijn bijna zeker niet ontstaan voor het vliegen. Ze ontstonden bij gevederde dinosauriërs, vermoedelijk voor warmte-isolatie of ten behoeve van seksueel pronkgedrag. Pas veel later, toen sommige diersoorten naar de boomtoppen klommen en daarvandaan glijvluchten begonnen te maken, kregen deze veren een herbestemming voor het vliegen. De middenoorbeentjes van zoogdieren (de hamer, het aambeeld en de stijgbeugel die het geluid doorgeven aan het binnenoor) zijn ooit ontstaan als botjes in de kaak van vroege voorlopers van zoogdieren. Toen de kaakconstructie veranderde, raakten die botjes hun oude functie kwijt en kregen ze een herbestemming in het gehoor. Longen zijn waarschijnlijk geëvolueerd uit gasuitwisselingsorganen bij vroege vissen die in zuurstofarm water leefden; pas later werden ze gebruikt voor ademen op het droge.
Doelloos, maar wel reconstrueerbaar
De kern van exaptatie is eenvoudig: een eigenschap is ouder dan de functie die ze nu vervult8; ze was er al voordat ze nuttig werd. Bij een adaptatie heeft de eigenschap zich juist verspreid omdát ze nuttig was: eenden met betere zwemvliezen bleken grotere overlevingskansen te hebben, dus werden zwemvliezen in de loop van vele generaties algemener. Bij een exaptatie bestond de eigenschap al om een andere reden, of zonder een bijzondere reden, en bleek ze pas later nuttig voor wat ze nu doet. Denk opnieuw aan de veren van vogels, die ontstonden bij dinosauriërs die niet konden vliegen en pas veel later bleken die veren bruikbaar voor een heel nieuwe functie: vliegen. De veer was er dus eerder dan het vliegen.
Zowel bij adaptatie als bij exaptatie is geen sprake van een plan of een bedoeling; de evolutie streeft namelijk nergens naar. Het onderscheid gaat dus niet over doelgericht versus doelloos, maar over de vraag: was een eigenschap al nuttig terwijl ze ontstond, of bestond ze al voordat ze nuttig werd? En hoe stel je dat vast, als er niemand bij was om het ontstaan te zien?
Het antwoord is dat je het reconstrueert uit bewijsmateriaal. Hierbij helpen drie soorten aanwijzingen. De eerste is de volgorde in de tijd: als een eigenschap al bij voorouders bestond, voordat ze haar huidige functie kreeg, dan is ze ouder dan die functie. Denk hier weer aan de veren bij gevederde dinosauriërs die niet konden vliegen. Zij bewijzen dat veren er eerder waren dan het vliegen en veren zijn dus een exaptatie. De tweede aanwijzing is de vorm zelf: een eigenschap die zich samen met haar functie heeft ontwikkeld, is daar meestal goed op afgestemd, terwijl een herbestemde eigenschap vaak sporen van haar oude functie draagt9, of waarvan zelfs de oude functie een raadsel blijft. De derde aanwijzing is de vergelijking tussen soorten: door te kijken welke verwanten het kenmerk wel en niet hebben, kun je reconstrueren wanneer en in welke vorm het oorspronkelijk ontstond.
Het onderscheid tussen adaptatie en exaptatie is dus geen kwestie van gissen naar bedoelingen, maar een empirische10 vraag, te beantwoorden via fossielen, stambomen en vergelijkend onderzoek. En precies dat maakt het ook toetsbaar.
Dan is er nog ‘integratie’
Eén begrip is hier nog onmisbaar, omdat het een kant belicht die in de hele discussie niet gemist mag worden: integratie. Dit is een functionele term die wijst op het verschijnsel dat losse onderdelen samen iets gaan doen wat ze afzonderlijk niet kunnen11. Hart, ademhaling, stem, gezicht, gehoor en sociale oriëntatie zijn elk afzonderlijk bestudeerd door verschillende vakgebieden. Maar bij zoogdieren werken ze als één samenhangend regulatiesysteem. Dit verschil in analyseniveau is precies wat Porges en de vergelijkend fysiologen langs elkaar heen doet praten.
Taylor en zijn collega’s bestuderen losse onderdelen: zenuwvezels, hersenstamkernen, geleidingssnelheden. Porges kijkt naar wat die onderdelen samen produceren. Beide benaderingen zijn van belang en legitiem; ze opereren alleen op verschillende niveaus, en wanneer ze met elkaar in debat treden alsof het om hetzelfde niveau gaat, ontstaan makkelijk misverstanden.
Een van die misverstanden kondigde ik eerder al aan, bij het woord “tak”. Wie de ventrale en de dorsale vagus voorstelt als twee aparte zenuwbanen, vertaalt een functioneel onderscheid ten onrechte in een fysieke scheiding. En precies zo'n verwarring van niveaus, het ene moment over losse vezels praten, het volgende over een samenhangend systeem, voedt een deel van de tegenstelling tussen Porges en zijn critici. Veel van wat als onverenigbaar wordt gepresenteerd, blijkt bij nader inzien een kwestie van praten op verschillende niveaus.
Porges’ eigen herformulering: een halve stap richting exaptatie
Een interessante ontwikkeling in dit debat is dat Porges zelf, sinds ongeveer 2021, in zijn formuleringen heel dicht bij het exaptatie-idee terecht is gekomen. Zonder de term te gebruiken, beweegt hij conceptueel wél in die richting.
In zijn artikel uit 2021 in het tijdschrift Comprehensive Psychoneuroendocrinology spreekt Porges niet meer over een ‘uniek zoogdier-systeem’, maar over een ‘herbestemd’ systeem (Porges, 2021). Het Engelse woord dat hij gebruikt is repurposed, wat letterlijk ‘opnieuw bestemd’ of (letterlijker vertaald) ‘opnieuw van een doel voorzien’ betekent. Strikt genomen klinkt daarin nog een bedoeling in door, alsof iets doelbewust een nieuwe taak krijgt, terwijl we weten dat exaptatie zonder plan plaatsvindt. Maar op die nuance na is de beweging die Porges maakt opmerkelijk dicht bij het exaptatie-idee.
Voorbeelden van zijn eigen formuleringen: “het zenuwstelsel kreeg een herbestemming om verdedigingsstrategieën te onderdrukken ten gunste van sociaal gedrag”. Elders schrijft hij dat de evolutie het zoogdier-vagale complex zou hebben herbestemd om sociaal gedrag te faciliteren. In zijn artikel uit 2022 in Frontiers in Integrative Neuroscience zegt hij het nog explicieter: “Evolutie is een proces van modificatie waarin bestaande structuren en circuits worden aangepast om adaptieve functies te vervullen.” (Porges, 2022). Dat laatste is bijna woordelijk wat Gould en Vrba in 1982 met exaptatie bedoelden. Daarmee erkent Porges in feite dat het bouwsteen-niveau niet houdbaar is, en verschuift hij naar het synthese-niveau. Maar hij benoemt dat niet als correctie van zijn eerdere claims.
Een tweede verschuiving betreft het anatomische referentiepunt. In zijn oorspronkelijke werk uit de jaren negentig sprak Porges over de NAmb, de hersenstamkern die wat meer naar voren ligt (oftewel ventraal) en waaruit de snellere vagale vezels ontspringen, als de bron van wat hij de ‘smart vagus’ noemde (Porges, 1995, 2011). In zijn werk vanaf 2021 spreekt hij consistent over het ventraal vagale complex, een grotere groep zenuwcellen die niet alleen de NAmb omvat, maar ook de hersenstamkernen die de gezichtspieren, de kaakspieren en de spiertjes in het middenoor aansturen (Porges, 2021, 2023).
Het zoogdier-eigene zit dus niet in één losse hersenkern, en evenmin in een grotere verzameling kernen op zich. Het zit in iets wat zoogdieren uniek maakt onder de gewervelden: een beweeglijk, expressief gezicht, en in de manier waarop oude vagus-bouwstenen daarmee zijn gaan samenwerken. De meeste zoogdieren hebben een echte gelaatsmusculatuur, de spieren onder de huid waarmee we glimlachen, fronsen en onze blik laten spreken. Vissen, reptielen en vogels missen die volledig; een hagedis kan niet fronsen, een vogel zijn gezicht niet vertrekken12. Dat nieuwe gezicht vraagt om aansturing, en het is die aansturing die bij zoogdieren verweven raakte met de oude regulatie van het hart.
Tegelijk laat Porges andere uitspraken staan die met deze halve stap niet zijn opgelost. Hij blijft schrijven over ‘de overgang van asociale reptielen naar sociale zoogdieren’ (Porges, 2021). Dat klopt empirisch niet helemaal. Onderzoekers als Doody, Burghardt en Dinets hebben in 2021 een heel boek gewijd aan het sociale leven van reptielen, met ouderzorg, groepsvorming en collectieve nestbescherming (Doody, Dinets, & Burghardt, 2021). In 2023 publiceerden ze een artikel in Biological Psychology, waarin ze de aanname dat reptielen asociaal zijn rechtstreeks weerleggen (Doody, Burghardt, & Dinets, 2023). De exaptatie-formulering die ik hier voorstel, lost dit spanningsveld op: ze laat sociaal gedrag bij reptielen volwaardig staan, zonder dat het zoogdier-eigen samenspel van gezicht, stem en hart daardoor wordt ontkend.
Een laatste kwestie betreft de wetenschappelijke status van de theorie zelf. In 2021 schreef Porges: “De theorie was niet bedoeld om bewezen of weerlegd te worden, maar om geïnformeerd te worden door onderzoek en gemodificeerd.” (Porges, 2021). Dat is in wetenschapsfilosofisch opzicht een problematische uitspraak, die Porges ook geregeld door critici is aangewreven. Een theorie die zich aan toetsing onttrekt (wat overigens niet is wat Porges zei noch bedoelde), plaatst zichzelf buiten de wetenschap13. Hier biedt de exaptatie-vorm van de polyvagaaltheorie een onverwachte uitweg. Ze maakt de theorie juist méér toetsbaar in plaats van minder. De centrale stelling verschuift naar de aanwezigheid en werking van een samenhangend gezicht-stem-hart-systeem bij zoogdieren, en weg van anatomische uitspraken die de vergelijkende fysiologie inmiddels heeft weerlegd. Wat overblijft, is empirisch te onderzoeken14.
De polyvagaaltheorie als theorie van exaptatie
Wanneer we het Taylor-artikel en de polyvagaaltheorie naast elkaar leggen en bekijken door de bril van exaptatie, lost de schijnbare tegenstelling op in een werkbare synthese.
Het is daarbij verhelderend te zien dat exaptatie hier op drie niveaus tegelijk werkt, en dat het zinvol is die niveaus uit elkaar te houden.
Op het evolutionaire niveau is exaptatie een vorm-en-functie duiding: oude bouwstenen krijgen een nieuwe rol in een nieuwe omgeving. Op het neurobiologische niveau zien we een reorganisatie: bestaande netwerken die anders gekoppeld raken, een andere hiërarchie krijgen en op nieuwe manieren met elkaar gaan communiceren. Op het functionele niveau zien we de integratie: hart, ademhaling, stem, gezicht en gehoor die als één samenhangend systeem gaan opereren.
Deze drie niveaus beschrijven niet drie verschillende verschijnselen, maar één verschijnsel vanuit drie perspectieven. Wie alleen op het niveau van de losse bouwstenen kijkt (zoals in de discussie soms gebeurt), ziet de continuïteit met andere gewervelden en concludeert dat er bij zoogdieren niets bijzonders aan de hand is. Wie alleen op het functionele niveau kijkt, ziet de bijzonderheid, maar mist de evolutionaire continuïteit. Pas wanneer de drie niveaus tegelijk in beeld blijven, valt het verhaal op zijn plek.
De bouwstenen die Porges aanvankelijk als typisch zoogdier-eigen presenteerde (gemyeliniseerde vagusvezels, een ventrale groep hart-aansturende vaguscellen, een ademritme dat de hartslag moduleert) zijn empirisch gezien geen uitvindingen van zoogdieren. Ze maken deel uit van een veel oudere uitrusting die alle gewervelden delen. Maar wat bij zoogdieren wél nieuw is, is de manier waarop deze oude bouwstenen in een samenhangend ensemble zijn geïntegreerd dat nergens anders in het dierenrijk in deze vorm bestaat.
Bij zoogdieren
Wat bij zoogdieren werkelijk nieuw is, zijn dus niet de losse bouwstenen, maar de combinatie. En deze combinatie laat zich het duidelijkst zien bij iets wat geen enkel ander gewerveld dier bezit: een beweeglijk, expressief gezicht. Dat gezicht zelf is wel een nieuwe anatomische verworvenheid; wat exaptatie heet, zit niet in dat gezicht op zich, maar in de manier waarop oude vagus-bouwstenen daarmee zijn gaan samenwerken.
Zoogdieren zijn de enige diergroep met een echte gelaatsmusculatuur, de spieren onder de huid waarmee we glimlachen, fronsen, knipperen en onze blik laten spreken. Die spieren zijn bij zoogdieren divers en gedifferentieerd en zijn verbonden met de bewegingen van lippen, kaak, tong en strottenhoofd. Vissen, reptielen en vogels missen dit volledig: een hagedis kan niet fronsen, een vogel zijn gezicht niet vertrekken. Daarbovenop ontwikkelt het strottenhoofd zich tot een verfijnd apparaat voor stem en intonatie, en krijgt het middenoor een regelsysteem dat geluidsfrequenties kan dempen of versterken, afgestemd op de geluiden die soortgenoten maken. Bij baby’s coördineren verschillende hersenzenuwen samen het zuigen, slikken en ademen, een patroon dat bij geen enkel reptiel of amfibie in deze vorm bestaat.
Dat dit alles bij zoogdieren mogelijk werd, hangt samen met een tweede kenmerk: warmbloedigheid. Die is overigens niet uniek voor zoogdieren. Vogels zijn ook warmbloedig, en er zijn sterke aanwijzingen dat de gemeenschappelijke voorouders van zoogdieren, vogels en krokodilachtigen een vorm van warmbloedigheid bezaten (Grigg et al., 2021). Een warmbloedig dier houdt zelf zijn lichaamstemperatuur op peil. Het kan zich daardoor iets permitteren wat een koudbloedig dier niet kan, namelijk een continue, fijngevoelige bijsturing van de fysiologische toestand, mede afhankelijk van wat er sociaal gebeurt. Die permanente bijsturing kost energie, en juist die energie heeft een warmbloedig dier beschikbaar. Warmbloedigheid is daarmee niet zelf de vernieuwing, maar een voorwaarde die de vernieuwing mogelijk maakt.
En hier wordt zichtbaar wat de exaptatie precies is. Niet de mimiek zelf, dat is een nieuwe verworvenheid. En niet de vagale rem op het hart; die is juist oeroud en delen wij met longvissen en reptielen. De exaptatie is de verbinding tussen die twee: een oud mechanisme, de snelle vagale rem die het hart van moment tot moment kan bijsturen, dat in dienst komt van iets volstrekt nieuws, namelijk een gezicht dat een innerlijke toestand kan tonen en bij een ander kan aflezen. De oude hartregulatie kreeg een sociale rol die pas mogelijk werd toen er een expressief gezicht was om mee te communiceren, en toen er genoeg energie was om die afstemming voortdurend te laten plaatsvinden. Dit is exaptatie in haar meest duidelijke vorm: een oud onderdeel dat, in een nieuwe configuratie, een nieuwe betekenis krijgt.
En in feite reikt die herbestemming verder dan deze ene koppeling van hart en het gezicht. Want het sociale engagementsysteem is opgebouwd uit een hele reeks oude onderdelen. De middenoorbeentjes waarmee we de stem van een ander verstaan, ontstonden, zoals we eerder zagen, als fragmenten van de kaakbotten van reptielen en haaien en kregen pas later hun rol in het gehoor. Het strottenhoofd, waarmee we stemklank en intonatie vormen, gaat terug op een klep die de lagere luchtweg afsluit, een functie die ze nog altijd vervult; de vorming van stem kwam daar pas later bij. En de vagale rem op het hart is honderden miljoenen jaren oud. Sommige van deze onderdelen waren bovendien al eerder met elkaar verbonden. Bij duikende reptielen werken larynx en hartslag samen om tijdens een duik zuurstof te sparen. De losse stukken, en zelfs sommige van hun verbindingen, zijn dus geen uitvinding van zoogdieren.
Wat zoogdieren onderscheidt, is iets anders. Rond één werkelijk nieuwe verworvenheid, het beweeglijke gezicht, kwam dit oude samenstel in dienst te staan van een nieuwe functie: niet het sparen van zuurstof of het beschermen van de luchtweg, maar het tonen, laten klinken en aflezen van een innerlijke toestand tussen soortgenoten. Het sociale verbindingssysteem is daarmee niet zomaar een voorbeeld van exaptatie. Het is een heel ensemble van oude onderdelen en oude verbindingen, herbestemd, uitgebreid met een nieuw element, en als geheel in dienst gesteld van iets wat geen van de delen afzonderlijk ooit kon: sociale verbinding.
Hoe strak dit geheel neuraal geïntegreerd is, in de zin die Porges voor ogen staat, is een vraag die nog verder onderzoek vergt; daarover doet de vergelijkende fysiologie van Taylor en zijn collega’s geen uitspraak. Maar dát zoogdieren beschikken over een communicatiekanaal dat geen ander dier heeft, en dat dit kanaal de oude hartregulatie heeft meegenomen in een nieuwe, sociale functie, staat buiten kijf.
En juist dat communicatiekanaal raakt aan wat het hart 😉 van de polyvagaaltheorie vormt: het begrip veiligheid. Want waar dit zuinige, fijngevoelige regulatiesysteem werkelijk verschil maakt, is in de nabijheid van soortgenoten. Een dier dat alleen kan kiezen tussen sympathische verdediging, vechten of vluchten, of dorsale shutdown, ervaart de nabijheid van een ander al snel als bedreiging. Maar een dier dat zijn hartfrequentie en zijn mimiek via de ventrale vagus subtiel kan bijsturen, kan in de buurt van een ander zijn zonder in de verdediging te schieten. Het kan nabijheid verdragen en zelfs opzoeken. Dat is fysiologisch belangrijk, want het betekent dat de nabijheid van een soortgenoot niet onmiddellijk in een vecht- of vluchtreactie hoeft om te slaan. En daar vinden we ook de voorwaarde voor alles wat daarna sociaal mogelijk werd: niet vechten maar afstemmen, niet vluchten maar verbinden, niet alleen overleven maar samen overleven. De ventrale vagus heeft die samenwerking niet in zijn eentje veroorzaakt, want samenwerking, wat uiteindelijk tot beschaving leidde, heeft vele wortels: van een verlengde kindertijd waarin meerdere volwassenen voor één kind zorgen, via gedeelde taal en intentionaliteit, tot het culturele vermogen om kennis door te geven aan volgende generaties. Maar de ventrale vagus leverde wel het lichamelijke fundament waarop een gevoel van veiligheid in elkaars nabijheid kon ontstaan, en zonder dat fundament was de rest niet mogelijk geweest. Een oud mechanisme, ooit werkzaam in de regulatie van het hart, werd zo de stille voorwaarde voor een leven onder en met soortgenoten.
Wat dit betekent voor theorie en praktijk
Deze herformulering heeft gevolgen die het waard zijn om expliciet te maken.
Voor de polyvagaaltheorie zelf betekent het exaptatie-perspectief dat de evolutionaire stellingen zoals Porges ze oorspronkelijk formuleerde niet meer als noodzakelijke onderbouwing kunnen dienen. De uitspraak dat alleen zoogdieren een gemyeliniseerde vagus zouden hebben, of dat de dorsale motorkern evolutionair ouder is dan de nucleus ambiguus, houdt geen stand bij wat de huidige vergelijkende fysiologie laat zien. Die uitspraken hoeven dan ook geen onderdeel meer te zijn van het theoretische bouwwerk. Wat overblijft, en wat empirisch en klinisch goed verdedigbaar is, is de stelling dat zoogdieren, en specifiek de mens, een samenhangend gezicht-stem-hart-systeem hebben dat sociaal gedrag en lichamelijke regulatie aan elkaar koppelt op een manier die elders in het dierenrijk geen evenknie heeft. Deze stelling staat of valt niet met de vraag of de nucleus ambiguus al bij hagedissen voorkomt.
Voor de klinische praktijk is dit een geruststellende uitkomst. De bruikbaarheid van polyvagaal-geïnformeerde benaderingen (sociaal contact als regulator van fysiologische toestand, vagale tonus als marker van flexibiliteit, co-regulatie als behandelelement) is grotendeels onafhankelijk van de vraag of de critici of Porges gelijk hebben over haaien en longvissen. De waarnemingen die de klinische theorie dragen, zijn waarnemingen aan mensen onder sociale en meer stressvolle omstandigheden. Die blijven geldig zolang ze in eigen recht empirisch worden bevestigd.
En toch raken het verre verleden en de spreekkamer elkaar. Wat zich over honderden miljoenen jaren aftekende, namelijk dat nabijheid pas mogelijk wordt wanneer het lichaam zich veilig genoeg voelt om de verdediging te laten zakken, is precies wat zich in het klein in elke behandelkamer afspeelt. Co-regulatie werkt niet in de eerste plaats door een techniek, maar doordat het zenuwstelsel van de ander de nabijheid opnieuw als veilig leert registreren. De therapeut doet, misschien wel in één ontmoeting, wat de ventrale vagus over de loop van de evolutie mogelijk maakte.
Voor de wetenschappelijke discussie tot slot betekent het exaptatie-perspectief dat de woordenstrijd tot rust mag komen. Taylor en zijn collega’s eindigen hun artikel met een opmerking die als een uitnodiging aan Porges gezien mag worden: een theorie verdient regelmatige toetsing, en zij zijn benieuwd hoe de polyvagaaltheorie zich onder die toetsing zal ontwikkelen.
De vraag is niet wie het gelijk aan zijn kant heeft. De vraag is of de polyvagaaltheorie bereid is haar woordenschat zo bij te stellen dat de bouwsteen-stelling losgemaakt wordt van de samenhang-stelling, en daarmee een evolutionair preciezere én klinisch sterkere formulering krijgt.
Hoe oud?
Wat is dan het antwoord op de vraag waar dit artikel mee begon: hoe oud is de ventrale vagus eigenlijk?
Dat hangt af van wat je ermee bedoelt. De anatomische bouwstenen, ventrolateraal gelegen cardiale vagale neuronen met gemyeliniseerde vezels naar het hart, zijn minstens vijfhonderd miljoen jaar oud en gaan terug tot de gemeenschappelijke voorouder van alle gewervelden. De respiratoir gemoduleerde versie ervan, het patroon waarin het hartritme schommelt op het ritme van de ademhaling, is ongeveer vierhonderd miljoen jaar oud en hangt samen met het ontstaan van longademhaling. De specifieke integratie van hart, ademhaling, stem en gezicht in één samenhangend regulatiesysteem rondom een beweeglijk en expressief gezicht is ongeveer tweehonderd miljoen jaar oud en valt samen met het ontstaan van de zoogdieren.
De ventrale vagus is dus tegelijk heel oud en heel “jong”. Welk antwoord klopt, hangt af van het niveau waarop je kijkt. En dat is waar het exaptatie-perspectief zijn waarde laat zien.
Tot besluit
De vergelijkende fysiologie van Taylor, Wang en Leite is geen aanval op de polyvagaaltheorie als therapeutisch denkkader. Het is een correctie van een specifieke evolutionaire claim, op een terrein waar deze drie onderzoekers tot de grootste autoriteiten ter wereld behoren. Wie hun werk leest als een afkeuring van de PVT, mist een kans op verheldering. Wie het leest als bouwmateriaal voor een rijpere formulering van de polyvagaaltheorie, een formulering die haar bouwstenen onderscheidt van haar samenhang en die samenhang verklaart als exaptatie in plaats van als anatomische uitvinding, heeft een denkraam in handen dat de klinische praktijk beter ondersteunt dan de oorspronkelijke versie deed.
Een afbakening is hier op haar plaats. De kritiek die in dit artikel is besproken, is de evolutionaire kritiek: de vraag of de bouwstenen van het systeem werkelijk zoogdier-eigen zijn. Dat is niet de enige kritiek die de polyvagaaltheorie heeft gekregen. Er zijn ook methodologische vragen, bijvoorbeeld over hoe ventrale vagale activiteit het beste kan worden gemeten, en er is discussie over de klinische claims. Die vallen buiten het bestek van dit artikel. Wat ik hier heb willen laten zien, is dat juist de evolutionaire bezwaren, die op het eerste gezicht het hardst lijken aan te komen, hun scherpte verliezen zodra je ze door de bril van exaptatie bekijkt. De bouwstenen zijn oud, dat punt scoren Taylor en collega’s. Wat zoogdieren ermee doen is nieuw; dat punt blijft voor Porges. En exaptatie is het sleutelpunt waarop die twee waarheden samenkomen.
Een korte methodologische opmerking tot slot. Eerder in dit artikel merkte ik op dat hart, stem en gezicht bij zoogdieren samen iets doen wat de losse delen niet kunnen, en ik beloofde daarop terug te komen. Wie toen aan emergentie dacht, schreef ik, had scherp opgelet. Emergentie is het verschijnsel dat een samenhangend systeem eigenschappen vertoont die in de losse onderdelen niet aanwezig zijn. Wie de polyvagaaltheorie als beschrijving van een geïntegreerd regulatiesysteem leest, ziet daarin een emergente laag: co-regulatie, gevoel van sociale veiligheid, prosodie en affectieve afstemming als eigenschappen die het hele systeem voortbrengt, niet de afzonderlijke onderdelen.
Eén vraag blijft tot slot hangen. Porges schoof de afgelopen jaren zelf al op richting een taal van herbestemming, van ‘repurposing’. Wanneer zou hij, gezien deze beweging, zelf de stap naar het begrip exaptatie hebben gezet?
Nabeschouwing
Het exaptatie-perspectief doet, als ik het overzie, drie dingen tegelijk in het polyvagale debat.
Ten eerste lost het een schijntegenstelling op die de discussie al jaren in een patstelling houdt: de keuze tussen zoogdier-uniciteit en evolutionaire continuïteit. Beide kanten hebben gelijk gehad, alleen op verschillende niveaus, en het exaptatie-begrip geeft de taal om dat verschil expliciet te maken.
Ten tweede verschuift het de zwaarste claim van de polyvagaaltheorie van een kwetsbaar anatomisch niveau, waar Taylor en zijn collega’s empirisch sterk staan, naar een synthese-niveau dat empirisch goed te verdedigen is. Wat bij zoogdieren werkelijk anders is georganiseerd, is niet de aanwezigheid van afzonderlijke bouwstenen, maar de integratie van gezicht, stem en hart in één samenhangend regulatiesysteem. Die stelling staat of valt niet met de vraag of bepaalde vagale vezels al bij de longvis voorkomen.
Ten derde, en dit lijkt paradoxaal maar dat is het niet, maakt het de polyvagaaltheorie juist toetsbaarder dan in haar oorspronkelijke vorm. De claims die overblijven gaan over wat in mensen meetbaar en onderzoekbaar is: de mate waarin gezicht, stem en hart op elkaar reageren, de werking van co-regulatie, de relatie tussen vagale tonus en sociaal gedrag. Dat zijn empirische vragen, geen evolutionaire claims die zich aan toetsing onttrekken.
Wat exaptatie niet doet, en daar wil ik eerlijk over zijn, is alle kritiek op de polyvagaaltheorie opheffen. Er liggen nog open methodologische vragen, zoals hoe ventrale vagale activiteit het beste kan worden gemeten. En de specifieke fylogenetische claims rond de evolutionaire herorganisatie van vagale circuits zijn plausibel, maar minder stevig onderbouwd dan het anatomische deel van de theorie. Hoe strak ook het integratiebeeld dat Porges schetst neuraal werkelijk gerealiseerd is, blijft een open empirische vraag.
Maar voor het evolutionaire deel van het debat, juist waar de PVT-gemeenschap tot nu toe het minst overtuigend heeft gereageerd, biedt exaptatie wat het debat nodig heeft: een herformulering waarin geen winnaar of verliezer wordt aangewezen, maar beide kanten op een hoger niveau worden verzoend. En het is niet zonder betekenis dat Porges zelf sinds 2021 al die kant op beweegt met zijn taal van “repurposing”. Hij gebruikt het woord exaptatie niet, maar hij zegt iets wat er heel dicht tegenaan ligt. Dit artikel maakt die beweging expliciet en plaatst ze in een evolutionair concept waar de biologie al sinds 1982 mee werkt.
Eén observatie tot slot. Misschien is de blijvendste bijdrage van deze hele oefening niet eens het exaptatie-argument zelf, maar de drielagenstructuur die mee komt: evolutionair, neurobiologisch en functioneel. Dat is geen exaptatie-technische kwestie meer, maar een denkraam dat ook in andere debatten in de polyvagaal-literatuur kan helpen om verwarringen tussen niveaus uit elkaar te halen. Als er iets uit dit stuk een eigen leven gaat leiden in de discussie, vermoed ik dat het misschien daar zit.
De oude vagus is ouder dan we dachten,
maar wat zoogdieren ermee deden, was nieuw.
Als je dit artikel lezenswaardig vond (ja toch, hoop ik?) en (nog) geen betaald abonnement wilt, mag je me ook trakteren op een cappuccino!
Geraadpleegde literatuur
Berta, A., Sumich, J. L., & Kovacs, K. M. (2015). Marine mammals: Evolutionary biology (3rd ed.). Academic Press / Elsevier.
Campbell, H. A., Leite, C. A. C., Wang, T., Skals, M., Abe, A. S., Egginton, S., Rantin, F. T., Bishop, C. M., & Taylor, E. W. (2006). Evidence for a respiratory component, similar to mammalian respiratory sinus arrhythmia, in the heart rate variability signal from the rattlesnake, Crotalus durissus terrificus. Journal of Experimental Biology, 209(14), 2628–2636. https://doi.org/10.1242/jeb.02278
Doody, J. S., Burghardt, G. M., & Dinets, V. (2023). The evolution of sociality and the polyvagal theory. Biological Psychology, 180, 108569. https://doi.org/10.1016/j.biopsycho.2023.108569
Doody, J. S., Dinets, V., & Burghardt, G. M. (2021). The Secret Social Lives of Reptiles. Baltimore: Johns Hopkins University Press.
Gould, S. J., & Vrba, E. S. (1982). Exaptation, a missing term in the science of form. Paleobiology, 8(1), 4–15.
https://doi.org/10.1017/S0094837300004310
Grigg, G., Nowack, J., Bicudo, J. E. P. W., Bal, N. C., Woodward, H. N., & Seymour, R. S. (2021). Whole-body endothermy: Ancient, homologous and widespread among the ancestors of mammals, birds and crocodylians. Biological Reviews, 97(2), 766–801.
https://doi.org/10.1111/brv.12822
Grossman, P. (2023). Fundamental challenges and likely refutations of the five basic premises of the polyvagal theory. Biological Psychology, 180, 108589. https://doi.org/10.1016/j.biopsycho.2023.108589
Grossman, P., & Taylor, E. W. (2007). Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biological Psychology, 74(2), 263–285. https://doi.org/10.1016/j.biopsycho.2005.11.014
Halder, G., Callaerts, P., & Gehring, W. J. (1995). Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science, 267(5205), 1788–1792.
https://doi.org/10.1126/science.7892602
Popper, K. R. (1959). The Logic of Scientific Discovery. London: Hutchinson.
Porges, S. W. (1995). Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A Polyvagal Theory. Psychophysiology, 32(4), 301–318.
https://doi.org/10.1111/j.1469-8986.1995.tb01213.x
Porges, S. W. (2011). The Polyvagal Theory: Neurophysiological foundations of emotions, attachment, communication, and self-regulation. New York: W.W. Norton.
Porges, S. W. (2021). Polyvagal Theory: A biobehavioral journey to sociality. Comprehensive Psychoneuroendocrinology, 7, 100069. https://doi.org/10.1016/j.cpnec.2021.100069
Porges, S. W. (2022). Polyvagal Theory: A science of safety. Frontiers in Integrative Neuroscience, 16, 871227.
https://doi.org/10.3389/fnint.2022.871227
Porges, S. W. (2023). The vagal paradox: A polyvagal solution. Comprehensive Psychoneuroendocrinology, 16, 100200. https://doi.org/10.1016/j.cpnec.2023.100200
Shubin, N. (2008). De vis in ons (M. van Nieuwstadt, Vert.). Nieuw Amsterdam.
Taylor, E. W., Wang, T., & Leite, C. A. C. (2022). An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the ‘Polyvagal Theory’. Biological Psychology, 172, 108382. https://doi.org/10.1016/j.biopsycho.2022.108382
Net als bij het woord “tak” verderop in de tekst, is “baan” hier een vereenvoudiging. Strikt genomen gaat het niet om een aparte zenuwbaan, maar om een bundel vezels met een eigen oorsprong en eigen eigenschappen, die meeloopt binnen de grotere bundel die de nervus vagus is. Ik gebruik “baan” hier omwille van de leesbaarheid, maar houd in gedachten dat het om vezels binnen één zenuw gaat, niet om een afzonderlijke kabel.
Taylors onderzoek omvat in werkelijkheid alle klassen van gewervelde dieren (van haaien tot zoogdieren) en daarnaast kreeftachtigen. De opsomming in de hoofdtekst beperkt zich tot de groepen die in dit artikel ter sprake komen. Universiteitsprofiel: https://www.birmingham.ac.uk/staff/profiles/biosciences/taylor-edwin.
Naast DMNX komen we ook geregeld de afkorting DVN tegen. Ze verwijzen naar dezelfde hersenstamkern, de dorso-mediale motorkern van de nervus vagus. DMNX is de afkorting zoals gebruikt in de polyvagale literatuur; DVN is de in de huidige vergelijkende fysiologie geaccepteerde afkorting (Taylor et al., 2022). Voor aansluiting bij Porges' terminologie hanteer ik in dit artikel DMNX.
Concreter: de afsplitsing tussen de longvis-lijn en de lijn die uiteindelijk tot zoogdieren zou leiden, ligt in het vroege Devoon, ongeveer 400 miljoen jaar geleden. De splitsing tussen de amfibie-lijn (waaruit padden voortkwamen) en de amniote-lijn (waaruit reptielen, vogels en zoogdieren ontstonden) is jonger, ongeveer 340 miljoen jaar geleden. Beide afsplitsingen liggen dus diep in het Paleozoïcum, ruim vóór het verschijnen van de eerste echte zoogdieren, wat rond 200 miljoen jaar geleden het geval was.
Strikt genomen ligt het iets ingewikkelder. Amfibieën hebben een hart met twee atria (= boezems) en één ventrikel (= kamer), waarbij zuurstofarm en zuurstofrijk bloed zich gedeeltelijk in dezelfde kamer mengen. De meeste reptielen (schildpadden, hagedissen, slangen) hebben ook één ventrikel, intern gedeeltelijk verdeeld door een onvolledige tussenwand. Wat veel mensen niet weten: bijna alle reptielen hebben niet één, maar twee aorta's, een linker en een rechter, naast de longslagader. Anders dan wij dus.
Krokodilachtigen vormen een fascinerende uitzondering binnen de reptielen. Zij hebben wél twee volledig gescheiden ventrikels, net als vogels en zoogdieren, maar bewaarden het reptielen-eigen twee-aorta-systeem. Bij hen ontspringt de rechter aorta uit de linker ventrikel (zoals te verwachten is; deze aorta heeft een vergelijkbare loop met die van de mens), maar de linker aorta ontspringt uit de rechter ventrikel (wat bijzonder is, maar dat gaat te diep voor dit artikel). Op het punt waar deze twee aorta’s elkaar passeren bij de basis van het hart, bevindt zich een opening die het foramen van Panizza heet. Daardoor kan bloed onder bepaalde omstandigheden alsnog tussen de twee aorta’s oversteken, en kan zuurstofarm bloed via de linker aorta naar het lichaam worden gestuurd zonder eerst door de longen te gaan. Dat is een prachtig evolutionair functioneel kenmerk: tijdens het duiken kunnen krokodillen hun longcirculatie effectief omleiden om zuurstof te sparen.
Voor het in dit artikel besproken mechanisme is het beslissende punt niet het aantal ventrikels, maar of bloed onder bepaalde omstandigheden kan worden omgeleid tussen long- en lichaamscirculatie. Bij vissen, amfibieën en bijna alle reptielen kan dat. Bij zoogdieren en vogels niet. Krokodillen laten in dat opzicht iets moois zien: ze tonen dat anatomische scheidingen en functionele scheidingen niet hetzelfde zijn. Twee ventrikels betekent niet automatisch dat de circulatie volledig gescheiden is.
Toen onderzoekers het eyeless-gen (en zijn zoogdier-tegenhanger Pax6) tot expressie brachten op een afwijkende plek bij een fruitvlieg, zoals op een poot, vleugel of antenne, groeide daar een compleet vliegenoog (Halder, Callaerts & Gehring, 1995). Het schakelgen gaf dus het startsein ‘bouw hier een oog’, en de vlieg voerde dat uit met haar eigen genetische gereedschap. Het effect is zo diep geconserveerd dat zelfs het Pax6-gen van een muis of een inktvis dit in een vlieg voor elkaar krijgt, ondanks dat deze diergroepen honderden miljoenen jaren geleden uiteen zijn geëvolueerd. De structuur van het oog is dus telkens opnieuw uitgevonden, maar het genetische gereedschap waarmee dat gebeurde is oeroud en gedeeld. Daarmee illustreert het oog op kleine schaal precies wat dit artikel op grotere schaal betoogt: oude bouwstenen die keer op keer opnieuw worden ingezet voor vergelijkbare doelen.
Het concept exaptatie is sinds het artikel van Stephen Jay Gould en Elisabeth Vrba in 1982 goed ingeburgerd in het evolutionaire denken. Een mogelijke tegenwerping is dat ik exaptatie in mijn artikel post-hoc inzet om een theorie te redden. Dat zou het concept miskennen: exaptatie is in de evolutiebiologie standaardgereedschap voor het begrijpen van functionele verschuivingen, geen ad-hoc reparatie.
Het is echt spitsroeden lopen met formuleren! Ik had eerst geschreven: “De kern van exaptatie is dat de huidige functie van een eigenschap niet de reden hoeft te zijn waarom die eigenschap ooit ontstond.” Een reden impliceert echter al snel een doel, en de hele evolutie heeft geen doel, geen vooropgezet plan. Ik zal vast op enkele plaatsen in het artikel niet kunnen ontkomen aan een onzuivere formulering, maar goed, de leesbaarheid van een artikel vraagt ook iets.
Dit tweede criterium, het herkennen van een eigenschap aan de sporen van haar verleden, is prachtig geïllustreerd door Stephen Jay Gould in zijn beroemde essay over de duim van de panda. De reuzenpanda heeft een “duim” waarmee hij bamboe vasthoudt, maar die duim is geen echte vinger. Het is een vergroot polsbeentje dat een nieuwe functie heeft gekregen. Juist het feit dat de constructie onhandig en geïmproviseerd overkomt, verraadt dat het niet voor het grijpen ontworpen is, maar een herbestemd botje dat toevallig van pas kwam. Gould gebruikte dit om te laten zien dat onvolkomenheid vaak een beter bewijs voor evolutie is dan perfectie: een perfect passende eigenschap zou immers ook bedacht kunnen zijn, maar een onhandige noodoplossing verraadt een geschiedenis van hergebruik. Bij de vagus zien we iets vergelijkbaars. Dat het ‘ademritmische’ fluctueren van de hartslag bij zoogdieren geen duidelijke fysiologische functie meer heeft, is precies zo’n spoor: het ziet eruit als een overblijfsel, niet als een doelgericht afgestemde aanpassing.
Empirisch betekent: gebaseerd op waarneming en ervaring, op wat je in de werkelijkheid kunt vaststellen, in plaats van op redenering, aanname of overtuiging alleen. Een empirische vraag is een vraag die je beantwoordt door te kijken, te meten en te vergelijken, niet door na te denken over wat logisch of wenselijk zou zijn. Het woord komt van het Griekse empeiria, dat ervaring betekent.
Wie nu aan het concept ‘emergentie’ denkt, heeft scherp opgelet; ik kom daar aan het slot op terug.
Walvissen en dolfijnen vormen hierbij een interessante uitzondering binnen de zoogdieren. Toen zij terugkeerden naar het water, hebben ze hun gelaatsmusculatuur grotendeels verloren, maar de nucleus ambiguus, de centrale kern van het ventraal vagale complex, hebben ze als zoogdier-erfgoed behouden. Hun sociale co-regulatie verloopt vooral via vocalisaties en lichaamscontact. Dat is anatomisch consistent met de gedachte dat de fylogenetische architectuur van het VVC behouden blijft, ook wanneer de specifieke uitwerking per zoogdiergroep verschilt (Berta, Sumich & Kovacs, 2015).
Porges’ formulering is ongelukkig gekozen en daardoor aanvalsgevoelig: critici lezen er niet-falsifieerbaar-willen-zijn in, vooral omdat vaak alleen het eerste deel van zijn uitspraak wordt geciteerd, dus “De theorie was niet bedoeld om bewezen of weerlegd te worden”! Als je de uitspraak in zijn geheel leest, dus: “De theorie was niet bedoeld om bewezen of weerlegd te worden, maar om geïnformeerd te worden door onderzoek en gemodificeerd”, dan lees je een verdedigbaardere intentie, namelijk dat zijn theorie eerder een breed organiserend kader is dan één enkele toetsbare hypothese. Dat onderscheid is in de wetenschapsfilosofie gangbaar: een overkoepelend raamwerk wordt zelden in zijn geheel bevestigd of weerlegd, maar levert wel toetsbare deelhypothesen op. Het ongelukkige zit hem dus niet in wat Porges vermoedelijk bedoelt, maar in hoe hij het formuleert.
Een toetsbare opzet zou kunnen zijn: laat twee mensen met elkaar in gesprek gaan en meet tegelijk hun gezichtsuitdrukkingen, de klank en melodie van hun stem, en hun hartslagvariabiliteit (HRV). Als gezicht, stem en hart bij zoogdieren werkelijk één regulatiesysteem vormen, zouden veranderingen in deze drie domeinen niet onafhankelijk van elkaar variëren, en zou een warmere stem en vriendelijker mimiek bij de één een meetbare verandering in de HRV van de ander oproepen, wat we co-regulatie noemen. Een manier om de claim te falsifiëren, loopt via mensen met het Möbius-syndroom, een aangeboren aandoening waarbij de gezichtsspieren niet werken: blijft bij hen de co-regulatie via stem en hartslag volledig intact, dan staat de stelling dat het gezicht een noodzakelijke schakel is, onder druk. Zo zijn er ook andere onderzoeken te bedenken.
Ik kreeg een bericht van Arent, dat ik met toestemming hier plaats en beantwoord:
Ik dacht nog aan (random) ‘mutatie’ vs adaptatie/exaptatie/emergentie waarbij de laatste term me deed denken aan breder systemisch denken… Mutaties en hierbij ook integraties zouden hierbij ‘random’/toevallig zijn, waarbij de species overleeft met de ‘meest handige/adaptieve/exaptieve’ mutatie(s)/integratie(s) van dat moment (in de omstandigheden van de wereld van dat moment). De vraag ‘wat hoort bij het ventrale ‘systeem’ zijn overigens ook je cellen, je benen, je gevoelens en gedachten .. toch? De rol van de epigenetica roept ook vraagtekens op (zijn mutaties helemaal random?)
Door het lezen van dit stuk (bravo!) gaat er inderdaad iets een eigen leven leiden haha.
Dat een aandoening in het middenoor (verkalking) waardoor geluid niet voldoende geleiding vind een exaptatie zou kunnen zijn….
Namelijk voor een leven lang in omgevingen/contexten zijn, waarin het aanhoren van mensen in de Fight respons mijn systeem teveel werd.
Hihi, kort door de bocht natuurlijk en een denkfout.
Desondanks heeft dit bij mij wel een glimlach gegenereerd op deze zondagochtend.
En het oorsuizen (dat ontstaat in specifieke onveilige situaties )kwam door dit artikel ook in een ander daglicht te staan. Dus persoonlijk ook iets aan gehad.
Bedankt voor het artikel. Succes 🍀